Известно, что Дарвин принял и объяснил знаменитый ракитник Адама (С. Adami) как прививочный гибрид. Морганисты с этим не согласны. С их точки зрения это периклинальная химера, в которой абсолютно независимо развиваются два рода тканей — внутренняя, принадлежащая L. vulgare, и внешняя — С. purpurens.
Возникает вопрос: если для С. Adami характерна такого рода автономность в развитии тканей разных родов, то чем объяснить (если исключить глубокое взаимовлияние) факт стерильности этого прививочного гибрида?
Известно, что половые органы у растений закладываются в субэпидермальном слое, который у С. Adami, с точки зрения ряда исследователей, принадлежит к чистому L. vulgare. Казалось бы, он должен беспрепятственно давать семена. Но семян, повторяем, С. Adami не образует. Причиной этого является уродливость пестика при нормальном развитии пыльников, несмотря на происхождение этих органов из одной зародышевой ткани.
На эти вопросы сторонники автономного развития частей химеры ответа не дадут. С нашей же точки зрения бесплодие, вызванное уродством пестика, является результатом взаимовлияния компонентов данного вегетативного гибрида.
Аналогичное явление наблюдается у вегетативного гибрида Crataego-mespilus. Со времён Баура общепринято считать, что внутренним компонентом здесь служит боярышник, а наружным — мушмула. Габерлянд(1926, 1927) провёл анатомическое изучение Crataego-mespilus и установил, что строение листьев, устьиц и сосудисто-волокнистых пучков у гибрида имеет промежуточный характер по сравнению с родительскими формами. Эти признаки не идентичны ни мушмуле, ни боярышнику.
Не только данные признаки поставили в тупик многих исследователей. Ещё более загадочно для морганистов ведут себя эпидермальные клетки верхней части листа. Оказалось, что эти клетки у Crataego-mespilus сходны с клетками боярышника. Они обладают прямыми стенками, мелкие, в то время как мушмула (по предположению она является внешним компонентом) имеет клетки удлинённой формы, с сильно волнистыми стенками (Weiss, 1925).
Особый интерес представляет и такая классическая химера, как Pyro-Cydonia, т. е. вегетативный гибрид между грушей и айвой, описанный в 1904 г. Люсьеном Даниелем.
По описанию Даниеля и детальному анатомическому исследованию Вейса (Weiss, 1930), во всех основных органах здесь ясно видны промежуточные признаки груши и айвы. Опушённость листьев здесь сильнее, чем у груши, и слабее, чем у айвы. Форма листа также промежуточная. Если груша имеет края листьев зубчатые, а айва — цельные, то у гибрида в верхней половине листа края зубчатые, а в нижней — цельные.
Строение эпидермальных клеток гибрида по своей величине и форме занимает серединное положение между клетками груши и айвы.
Н. П. Кренке, много работавший по вопросам трансплантации, также столкнулся с фактами взаимовлияний, в частности, у плодов от прививок S. lycopersicum и S. memphiticum.
«В полных гаплохламидных, цитологически (по хромосомам) проверенных плодах, — пишет Кренке, — антоциан, нормально присущий только покровному химерсимбионту Solanum memphiticum Mart., совершенно достоверно обнаружен во всех клетках мякоти томатной части химерных плодов… Все данные показывают, что в плодах химер, а вероятно и в других органах, налицо возможность специфического (подчёркнуто нами. — И. Г.) взаимодействия между химерсимбионтами» (Krenke, 1933).
И в последующих разделах упомянутой работы Кренке приводит, хотя и с оговорками, интересный случай изменчивости эпидермиса у химеры S. lycopersicum-memphiticum. На рисунке 69, заимствованном из работы Кренке, представлены участки эпидермиса химерных плодов. Зрелые плоды S. lycopersicum(S. 1.) и S. memphiticum (S. m.) имеют резко отличный эпидермис. S. m. I— эпидермис S. memphiticum в гаплохламидной химере, a S. т. II — тот же эпидермис в диплохламидной химере. Из рисунка видно, что в положении однослойного покрова химеры S. т. I эпидермис S. memphiticum имеет явно утолщённые оболочки клеток и с уменьшенным количеством видимых пор. Менее ярко, но также заметна изменчивость эпидермальных клеток у диплохламидной химеры S. т. II. Изменчивость как в первом, так и во втором случае идёт в сторону второго (внутреннего) компонента — томата.
Такова фактическая сторона вопроса о независимости растительных тканей при прививках.
Рассмотрим ещё одну группу фактов, служащих часто аргументом против вегетативной гибридизации. Мы имеем в виду такие случаи, когда при посеве семян из плодов химерных растений получались чистые представители или одного (внутреннего) или другого (внешнего) компонентов.
Что здесь можно сказать? Таких фактов очень немного. Большинство известных так называемых классических химер совершенно не дают семян, они стерильны.
Гильдебранду удалось в 1904 г. получить у С. Adami два плодика и в дальнейшие годы ещё несколько плодиков. Плоды имели всего по одному семечку. Сеянцы этих плодов в раннем периоде по цвету зелени и по числу долей листа (молодых листьев) напоминали С. Ada-mi, во взрослом состоянии они приближались к С. laburnum (Hildebrand, 1908). Следующий известный нам случай получения семенных потомств— у химер Винклера. Из пяти основных типов семена дали только три; S. tubingense и S. Gaertnerianum дали сеянцы, не отличимые от паслёна; S. proteus дал потомство томатного типа.
Остановимся несколько подробнее на характеристике плодоношения химерных растений Винклера.
Напомним, что растения S. tubingense вообще мало чем отличались от паслёна, за исключением более грубого опушения, что характерно для томата. При таком преобладании признаков одного из компонентов трудно было и ожидать заметных изменений плодов, хотя и в этом случае, как пишет Винклер, образовалось значительное количество партенокарпических плодов.
Здесь необходимо принять во внимание ещё и то, что химерным растениям свойственно давать адвентивные побеги, сходные по внешним морфологическим признакам или с чистым паслёном или томатом. Это наблюдалось и в наших опытах. В своей работе Винклер (1910) не указывает, с химерных или с адвентивных побегов получено семенное потомство. Если для этой цели брались плоды с побегов паслёнового типа, то. наиболее вероятно, что таким же должно быть и семенное потомство.
У S. Gaertnerianum цветение было нормальным, но цветки опадали, и только в конце лета изредка завязывались плоды. Они были гораздо крупнее паслёновых и окраску имели сине-фиолетовую, но при переходе от зелёной (незрелые плоды) к фиолетовой (полное созревание) проходили стадию жёлтой окраски, чего не бывает у паслёна и что характерно для томатов. Плоды опытного растения отличались от паслёна также сильно развитой плацентой и многокамерностью.
Склонность к увеличению камерности отсутствует у паслёна и, наоборот, характерна для томата. Семена полученных плодов легко и быстро прорастают, что является характерным для томата, но не для паслёна. Семена паслёна прорастают медленно, и для ускорения их прорастания часто необходимо прибегать к механическому повреждению семенных оболочек.
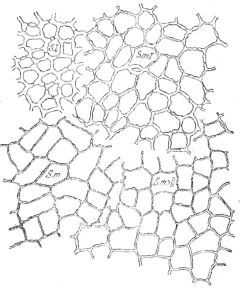
Рис. 69. Характер изменчивости эпидермиса у химер (по Кренке).
Плоды S. proteus ближе к томатным, имеют красную окраску, НО в 2–3 раза меньше. Кроме того, во многих клетках мякоти образовывался темносиний пигмент, отсутствующий у томата. Эпидермис плода также имел фиолетовые полосы и точки. Семена сходны с томатными, но гораздо мельче.
Аналогичная картина изменчивости наблюдалась у плодов S. Darwinianum, которых было получено много. Они имели красную с синим оттенком окраску, по величине же лишь немного крупнее паслёна. Плоды опадали до наступления полного созревания, и окраска доходила у них в лёжке. По внутреннему строению они напоминали больше томат, чем паслён. Семена во всех плодах оказались нежизнеспособными, и о семенных потомствах здесь вообще ничего нельзя сказать.
Растение S. Koelreuterianum также обильно цвело, но ни одного плода за все годы не дало. Между тем, согласно описанию, эта форма является монохламидной переклинальной химерой, т. е. субэпидермальный слой её состоит из чисто томатных клеток, и теоретически она должна была бы дать чистое томатное потомство.