
Рис. 64. Соматические пластинки: сверху вниз томата Гумберт (24 хромосомы); чёрного паслёна (72 хромосомы); вегетативного гибрида Гумберт/ чёрный паслён (26 хромосом).
В комбинации Гумберт/паслён чёрный соматические хромосомы и по числу и по морфологии отличались от материнской формы. Резко бросается в глаза их укороченность по сравнению с хромосомами контрольного томата: они почти вдвое короче. Наиболее часто встречающееся число их равно 26, т. е. имеется одна пара добавочных хромосом (рис. 64, внизу). Эта добавочная пара наиболее коротка, она отличается ещё и тем, что обычно располагается в несколько другой плоскости, чем остальные хромосомы. От встречающихся в чистых сортах томата добавочных хромосом она отличается тем, что скорее напоминает собою фрагменты, а не целые типичные хромосомы. Клетки с 24 хромосомами у этого гибрида встречаются как исключение. Изредка наблюдались пластинки и с меньшим числом, чем исходное 24, чего в чистых видах наблюдать не приходилось. В соматических клетках второй комбинации межродовых гибридов Гумберт/Деликатес метафазная пластинка мало отличается от чистого Гумберта. Правда, и здесь отличием было присутствие в некоторых клетках этого гибрида пары фрагментов, напоминающих тринадцатую пару предыдущего гибрида.
Редукционное деление у родительских видов (Гумберт и паслён) протекает нормально. У паслёна в метафазах первого и второго делений легко насчитать 36 хромосом, более мелких, чем у томата.
В редукционном делении обоих гибридов наблюдался ряд отклонений и ненормальностей. Особенно значительное и частое нарушение хода редукционного деления даёт комбинация Гумберт/ баклажан. Прежде всего сильно нарушается синхронность в прохождении фаз деления отдельными материнскими клетками. Независимость отдельных клеток выражена так резко, что присутствие в одном и том же пыльнике клеток самых начальных фаз деления и рядом уже готовых молодых пыльцевых зёрен не является редкостью. На ранних стадиях в пыльниках незаметно особых отклонений от нормы. Археспориальная ткань и мейотические профазы также не обнаруживают изменений. Но уже в метафазной пластинке, вместо гаплоидного числа 12, можно наблюдать большие числа хромосом — до 18 (рис. 65, 2 и 66, 2). Очевидно, наряду с бивалентами, здесь присутствует и некоторое число унивалентных хромосом, что указывает на преждевременное расхождение некоторых пар или, может быть, вообще на отсутствие у них конъюгации. В анафазах, как правило, наблюдается недружное расхождение хромосом: в то время как одни уже дошли до полюсов, другие находятся на пути, а третьи ещё лежат в экваториальной плоскости. Неправильное рассеивание хромосом по веретену и задержка части их ведёт к тому, что при образовании дочерних ядер часть хромосом не успевает включиться в них и остаётся в плазме в виде микроядер Микроядра или сливаются с основным ядром или долго сохраняются в плазме в виде хроматиновых включений, постепенно дегенерируя. Наблюдались и «незавершённые анафазы», когда в результате первого деления весь хромосомный комплекс клетки включается в одно, так называемое реституционное ядро, принимающее при этом характерную форму песочных часов (рис. 65, 2). В других случаях группы хромосом образуют самостоятельные ядра, и в клетке, прошедшей первое деление, оказывается больше двух ядер обычно резко разных размеров (рис. 65, 4). Так как те же аномалии могут происходить и во втором делении, то после двух делений материнская клетка может содержать вместо четырёх ядер большее их число (5, 6, 7) причём ядра могут быть разной величины. Среди плазмы встречаются отдельные хромосомы или сгустки хроматина, представляющие собой дегенерирующие остатки не включённых в ядра хромосом. Расстройство механизма деления на этой стадии выражается ещё одним явлением, никогда не встречающимся у чистых исходных форм, — слиянием дочерних ядер. Отдельные моменты слияния, наблюдавшиеся на препаратах, не позволяют сомневаться в его реальности. Сливается или одна пара ядер из четырёх или обе пары, давая в результате трёхъядерную или снова двухъядерную клетку (рис. 66, 3). Наконец, и все четыре дочерних ядра могут слиться и дать клетку с одним гигантским ядром. В связи с этим, наряду с молодыми, нормальными пыльцевыми зёрнами, в пыльниках можно найти гигантские клетки одноядерные, двухъядерные или клетки с четырьмя, плотно прилегающими друг к другу ядрами (рис. 66, 4), а также некоторое количество отмирающих клеток и пустых клеточных оболочек.
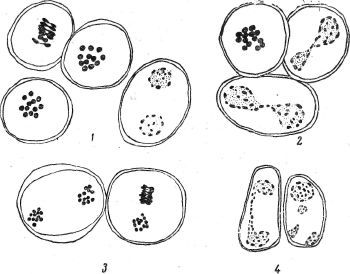
Рис. 65. Мейозис у вегетативного гибрида Гумберт/баклажан: 1 и 3 — нормальные картины I и II деления; 2 — пластинка с увеличенным числом хромосом, две клетки с «незавершёнными анафазами»; 4 — интерфаза ненормального мейозиса. Вместо диад — реституционное ядро в первой клетке, микроядра — во второй клетке.
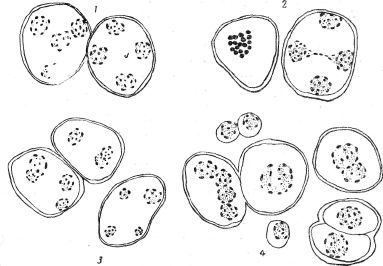
Рис. 66. Мейозис у вегетативного гибрида Гумберт/баклажан. 1 — нормальные тетрады; 2 — пластинка с увеличенным числом хромосом; рядом клетка с двумя ядрами и одним реституционным ядром; 3 — неровные ядра на стадии тетрад; картины слияния ядер; 4 — стадия молодой пыльцы. Наряду с нормальными пыльцевыми зёрнами, видны диадные клетки, гигантская монада, а также клетки со сливающимися ядрами.
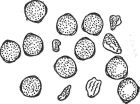
Рис. 67. Зрелая пыльца вегетативного гибрида Гумберт/баклажан.
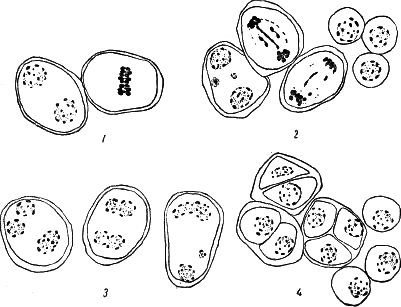
Рис. 68. Мейозис у вегетативного гибрида Гумберт/S. nigrum: 1 — нормальные материнские клетки пыльцы; 2 — отстающие хромосомы, микроядра и I интерфаза; 3 — II интерфаза; слияние ядер; 4 — стадия тетрад. Видны диады.
После распадения тетрады отдельные молодые пыльцевые зёрна сначала имеют вид хорошо развитых, густо наполненных плазмой клеток. Но с возрастом в гибридной пыльце происходят изменения. В некоторых пыльцевых зёрнах появляется потемнение, грубая зернистость, плазма делается губчатой, вакуолизированной, начинаются явления дегенерации. Пыльцевое зерно всё больше обедняется плазмой, становится пустым, наконец, наступает сморщивание, деформация и отмирание его (рис. 67).
Все описанные проявления расстройства механизма редукционного деления, только в меньшей степени, найдены были и у комбинации Гумберт/паслён (рис. 68). Необходимо отметить, что в обеих комбинациях, наряду с ненормальностями, наблюдались многочисленные картины совершенно типичного редукционного деления, не отличающиеся от контрольных форм. Нарушения редукционного деления не ведут в нашем случае к стерильности, так как в большей части материнских клеток оно протекает нормально.
1. У растений межсортовых и межвидовых вегетативных гибридов уклонений от родительских форм не обнаружено.
Объясняется это тем, что сорта и даже отдельные виды томатов близки между собою. Об этом свидетельствует лёгкость получения у томатов межвидовых половых гибридов.
2. Резкие уклонения от нормы наблюдаются при межродовых прививках. Эти уклонения выразились: во-первых, в нарушениях процесса редукционного деления и, во-вторых, в изменений количества хромосом в половых и соматических клетках.
3. Причину цитологических нарушений у отдалённых прививочных гибридов можно усматривать в следующем. Поскольку на основании современных данных признаётся, что репродукция хромосом происходит в результате образования дочерних хромосом de novo, то различия в пластических веществах прививки могут привести к качественным различиям хромосом данного комплекса.
4. Соответствующие изменения в хромосомном комплексе могут быть лишь одним из морфологических показателей более тонких изменений во всей структуре клетки.