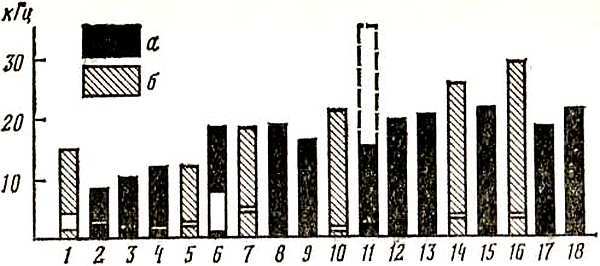
Как показывает рис. 10, верхний порог слуха птиц колеблется у разных видов от 10–12 кГц (голуби, куриные, пластинчатоклювые, дневные хищники) до 16–20 кГц (попугаи, воробьиные, совы, дятлы), однако в сезоны размножения воробьиные птицы способны реагировать на частоты до 35 кГц. Так, американские ученые Фрингс и Кук, используя электрооборонительные методики, установили, что именно до таких частот расширяются верхние границы слуха у скворца в весенние месяцы, тогда как в октябре они снижались до 16 кГц. Пороги слухового восприятия у волнистого попугайчика, определенные в 1940 г. Киехтом, составили 40–16 000 Гц. В этих же экспериментах Киехт определил и другой важный параметр — дифференциальные пороги, которые у волнистого попугайчика оказались близкими к человеческим и составили в области частот 0,3–1 кГц всего 0,3–0,7 % (2–5 Гц), тогда как у кур, например, они оказались равными при частоте 0,3 кГц 3 % (9 Гц), при 1 кГц — 2 % (20 Гц). Голуби же различают только 6 % частоты (в диапазоне 0,3–1 кГц). Значительно более суженной по сравнению с общим диапазоном слухового восприятия является зона повышенной слуховой чувствительности. Как видно из рис. 10, эта зона смещена или в низкочастотную область (голуби, куриные), или занимает область средних частот (воробьиные, попугаи), тогда как у сов она располагается в относительно высоких частотах (до 6 кГц и выше). Именно в области повышенного слуха располагаются самые низкие пороги звукового восприятия, как это показано на рис. 10. Как видно из этого рисунка, кривая слуховой чувствительности волнистого попугайчика несколько отличается от пороговой кривой человеческого слуха, хотя эти различия не слишком значительные.
Изучая характеристические частоты слуховых нейронов продолговатого мозга волнистого попугайчика, профессор Рурского университета И. Шварцкопф установил, что они стимулируются частотами 0,15-4 кГц, причем большая их часть хорошо реагирует на частоты в области 1,4 кГц.
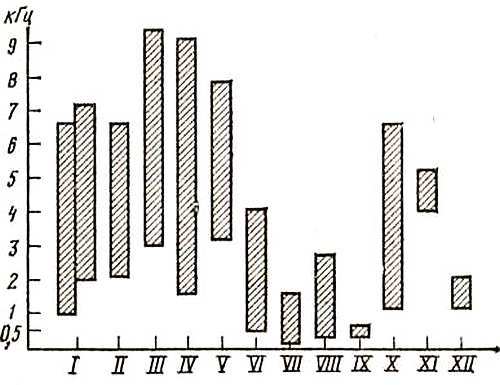
Если мы теперь обратимся к спектральному диапазону голоса птиц, то обнаружим лишь частичное совпадение его с частотными характеристиками слуха. На рис. 11 изображены общий диапазон голоса и область максимальных частот ряда видов птиц, слуховые возможности которых мы характеризовали выше. Сопоставляя рис. 11 с рис. 10, мы видим, что в большинстве случаев голосовой спектр (нередко и максимальные частоты) укладывается в область повышенного слуха или располагается близко от него. Так, например, крики волнистого попугайчика, охватывающие спектр 1,2–6,4 кГц, вполне укладываются? область его слухового восприятия, голоса сизого голубя и домашней курицы совпадают с областью повышенного слуха. В то же время обращает на себя внимание и значительно избыточная зона слухового восприятия, которую не «занимают» голоса этих видов. Например, слуховой диапазон ушастой совы лишь частично заполнен голосами слетков и взрослых птиц, тогда как большая его часть остается свободной. Как показали исследования одного из авторов этой книги, эта свободная зона, включающая область повышенного слуха, соответствует спектру жизненно важных для совы звуков — писков и шорохов мелких грызунов — полевок и мышей, на которых сова охотится, лоцируя их с помощью слуха.
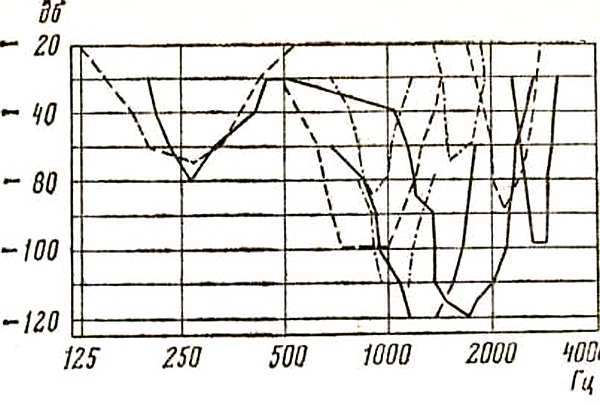
Профессор из ФРГ Иоганн Шварцкопф показал, что рецептивные зоны слуховых нейронов продолговатого мозга располагаются «внутри» диапазона голоса и некоторые из них совпадают с зоной повышенного слуха (рис. 12).
Таким образом, отсутствие жесткого соответствия между слухом и собственным голосом обеспечивает резерв, необходимый для восприятия разнообразной и сложной звуковой среды, в первую очередь голосов своих биоценотических партнеров — врагов или жертв, обнаружение и отслеживание которых в пространстве осуществляется с помощью слуха.
Характерно, что и голос человека, который в домашних условиях становится для попугая важнейшим «биоценотическим партнером», обеспечивающим его пищей, водой и этологическим комфортом, также совпадает с зоной повышенного слуха и это совпадение, безусловно, способствует птичьему «говорению».
И проще и сложнее
Итак, сопоставляя слуховые системы птиц и млекопитающих, вы обнаружите, что при сохранении общей схемы — наружное среднее, внутреннее ухо, слуховые ядра продолговатого, среднего, промежуточного мозга, больших полушарий — птицы не имеют наружной раковины, барабанная перепонка их велика и выпукла, в среднем ухе хрящевая пластинка и единственная косточка, единственная мышца, улитка втрое короче, слуховой коры нет и т. д.
С формально анатомической точки зрения слуховая система птиц выглядит проще. И в прошлом — это основание для того, чтобы ставить ее ниже на ступеньку по сравнению со слуховой системой млекопитающих. Длительное время среди анатомов она рассматривалась как эволюционный предшественник слуховой системы млекопитающих, поскольку она проще по строению и значит, по-видимому, хуже работает. И далее. Из этих представлений следовал и еще один вывод, уже организационный, — давайте не будем изучать слуховую систему птиц, пока не изучим вершины эволюции — слуховую систему млекопитающих. Вот почему слуховая система птиц оказалась значительно менее изученной по сравнению с таковой млекопитающих.
Но простота бывает разная. Бывает простота от примитивности, бывает простота от совершенства. Бывает простота, когда сложнейшие процессы осуществляются технологически совершенными, но внешне простыми устройствами.
И в самом деле, почему птицы должны копировать слуховую систему млекопитающих?
Мы говорили о происхождении птиц — они имеют других предков внутри группы рептилий и развивались параллельно с млекопитающими. Они ведут открытый образ жизни и летают. А это требует хорошего слуха, мгновенной ориентации в пространстве, где столько неприятностей, где так важно вовремя предостеречь партнера, выявить и установить местоположение врага. А полет требует экономии во всем, в первую очередь в размерах и весе. Поэтому эволюция птиц — это постоянное лавирование, поиск оптимального решения между противоположными тенденциями.
Отбор открывает «зеленую улицу» наиболее экономным, технологически простым, но надежным функционально эффективным решениям. Так появляются структурно упрощенные уши, подчас незаметные в оперении, скрытые обтекаемой формой тела, непривычные для наших глаз (привыкших к ушам-раструбам лошади или собаки) совиные уши на плоском круглом «лице» птицы.
По-видимому, мы должны отказаться от привычной идеи о том, что слуховая система птиц примитивна, что она эволюционный предшественник слуховой системы млекопитающих. По-видимому, правильнее рассматривать обе системы как независимо возникшие, параллельно развивающиеся, одинаково успешно обслуживающие тех, кто их использует.