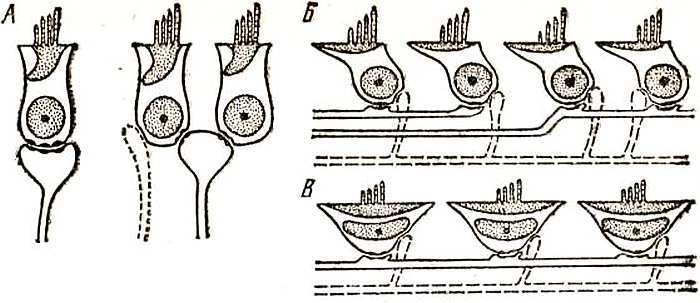
Для того чтобы доказать рычажную подвижность у птиц, пришлось пускаться на различные ухищрения. Перерезать хрящевую экстраколумеллю, с помощью которой косточка связана с барабанной перепонкой. Экстраколумелля имеет вид треноги, одна из ног которой упирается в центр перепонки и натягивает ее (вот почему перепонка у птиц выпукла, а не вогнута, как у млекопитающих), две другие располагаются в контакте с костным краем перепонки. Косточка прирастает к той точке экстраколумелли, где сходятся все три ее ноги.
Используя в качестве индикатора биоэлектрическую активность рецепторного отдела, вызванную действием звукового щелчка (кохлеарные потенциалы), и перерезая на разных уровнях опорные отростки — ноги экстраколумелли, можно получить чисто поршневой или чисто рычажный характер движений столбика и исследовать их роль в передаче звука раздельно. Опыты показали, что значение рычажной подвижности слухового столбика в работе слуховой системы птиц велико.
Сотрудник Московского университета В. Д. Анисимов разработал интересную методику изучения звукопередающей системы птиц — методику светящейся точки.
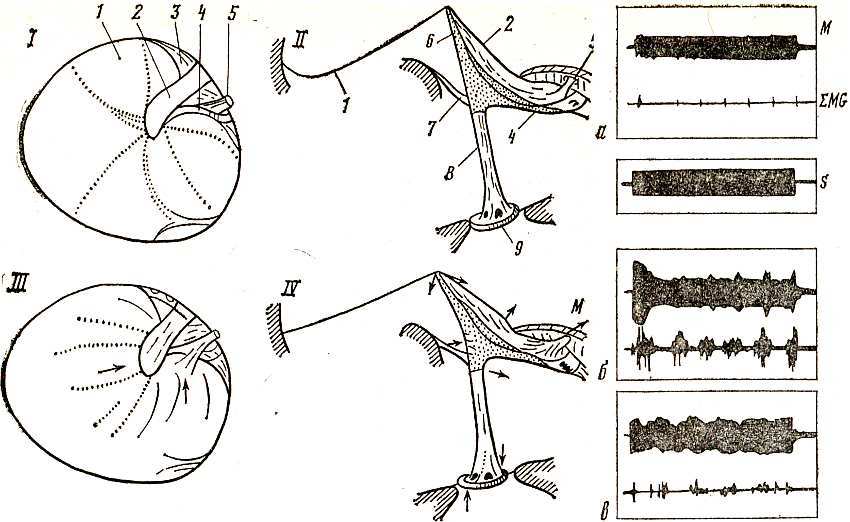
Наклеивая на различные участки звукопередающей системы кусочки блестящей фольги, отражающей свет, он зарегистрировал положение слуховой косточки и хрящевой экстраколумелли в различных динамических состояниях.
Другой важной методикой, разработанной В. Д. Анисимовым, было макетирование звукопередающей системы и ее функций на увеличенной кинематической модели, выполненной из прозрачного плексигласа. Задавая различные режимы сокращения среднеушной мышцы и вызываемого ею натяжения барабанной перепонки, можно было проследить характер подвижности звукопередающей системы, рычажные движения слухового столбика и экстраколумелли.
Напыление кристаллического серебра на различные элементы среднего уха, их подкрашивание и маркировка позволили заснять на пленку весь процесс движений, в том числе и рычажных звукопередающей системы. Эти же процессы повторились на увеличенной, модели среднего уха птиц, пропорционально увеличенной во всех звеньях.
Таким образом, было доказано, что среднее ухо птиц, иначе, чем у млекопитающих, устроенное, работает по тем же законам и решает аналогичные задачи.
И улитка другая
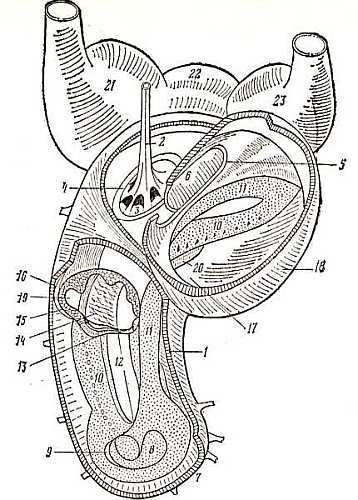
Еще в начале прошлого века биологи спорили (в этих спорах участвовал и великий Кювье) — есть или нет улитка у птиц. Так отличается она от улитки млекопитающих. Укороченная, расширенная, напоминающая сардельку (рис. 6), половина которой занята широкими затянутыми перепонкой окнами — овальным и круглым, она очень мало походила на привычную спираль улитки млекопитающих. И внутреннее строение ее также было необычным (рис. 7). Вестибулярный проток почти отсутствует, вытесненный рыхлой сосудистой покрышкой. Под покрышкой эндолимфа — вязкая жидкость, заполняющая кохлеарный проток, дно которого представлено базилярной мембраной с опорными и рецепторными клетками, прикрытыми покровной мембраной. У млекопитающих такой покрышки нет и оба протока разделены реснеровой мембраной. На базилярной мембране сложнейшее образование — кортиев орган с рецепторными клетками двух типов — наружными и внутренними, бесклеточным пространством между ними — туннелем, разнообразными опорными клетками. Каждая рецепторная клетка с выростами на верхнем конце — «волосками» (числом до 60), отсюда название клетки — волосковая. На нижнем конце клетки — контакты с отростками нейронов слухового нерва.
Рецепторные клетки внутреннего уха птиц в отличие от млекопитающих имеют другие модификации, так же как и опорные. Исследования Т. Б. Голубевой, Г. В. Ямаловой и Л. И. Прокофьевой выявили три такие модификации — высокие волосковые клетки, скошенные клетки и низкие волосковые клетки, отличающиеся формой, размерами, характером иннервации, деталями строения (рис. 8). Общее число волосковых клеток у вороны составляет около 14 000, у голубя значительно меньше (около 9000). В улитке птиц нет туннеля и вообще расположение рецепторных клеток представлено плотной упаковкой с многими рядами, продольными и поперечными (у млекопитающих только продольные ряды — один ряд внутренних волосковых клеток и три ряда — наружных). Кроме обычных прямых волосков — стереоцилий, присущих и млекопитающим, рецепторная клетка птиц песет по одному извитому волоску особого строения, который называется киноцилией.
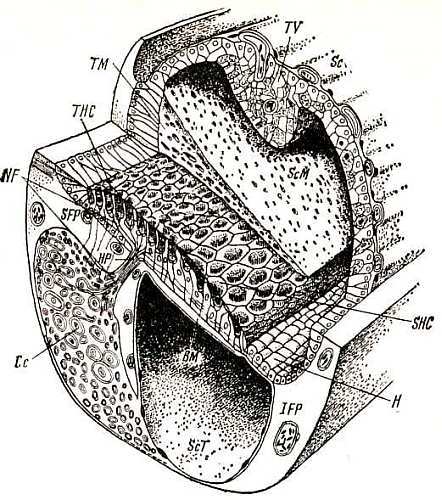
Базилярная мембрана птиц подложена широкими пластинами хрящей, оставляющими свободной только самую центральную ее часть в виде узкой продольной полоски. При этом длина базилярной мембраны и длина рецепторного эпителия в улитке серой вороны значительно превышают таковые голубя (соответственно 4,4 и 3,3 мм). Конец улитки заполнен особым образованием — лагеной. Лагена представлена группой волосковых клеток, прикрытых студенистой макулой с кристаллическими образованиями. Внешне это напоминает рецепторы вестибулярного аппарата и эту функцию лагене долго приписывали. В последние годы, однако, выяснилось, что нервные волокна, идущие от лагены, направляются далее в слуховые ядра мозга (как и в вестибулярные), и, следовательно, лагена может воспринимать звуки. У млекопитающих улитка не имеет лагены вовсе.