У фенотипической пластичности есть один аспект, который не до конца понимают даже некоторые биологи. Речь идет о том, что ее объем в той или иной степени тоже является мишенью отбора. Рассмотрим классический пример. Форма листьев водяного лютика зависит оттого, где он растет. Один и тот же генотип дает тонко рассеченные листья, если растение находится в воде, и широкие лопастные листья, если оно растет на суше. Если в воду погружена только нижняя часть водяного лютика, на нем образуются листья обоих типов. Более того, лист, сформировавшийся на границе между воздухом и водой, имеет широкую верхнюю и рассеченную нижнюю часть.
Наконец, говоря об эволюции путем естественного отбора, необходимо осознавать разницу между непосредственными причинами, определяющими то, как функционирует структура или процесс, и первопричинами, определяющими то, почему структура или процесс вообще существует. Вернемся к птицам, сменившим дубы на сосны.
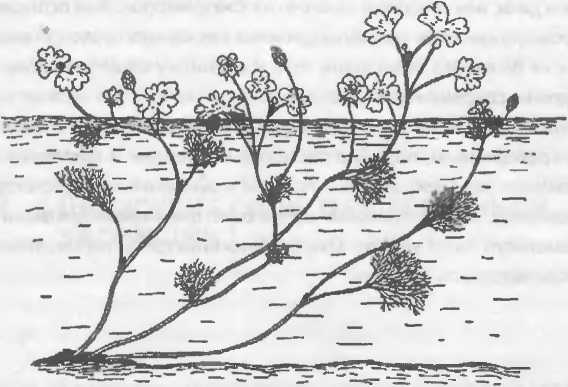
РИС. 17-2. Водяной лютик
(Ranunculus aquatilis)
обладает ярко выраженной фенотипической пластичностью — форма листа зависит от его положения. (Источник: Theodosius Dobzhansky, Evolution. Genetics, and Man [New York: Wiley, 1955}.)
Непосредственная причина микроэволюции — наличие аллеля Ь. Он предрасполагает к выбору сосен, а если точнее, предписывает определенное развитие эндокринной и нервной систем, опосредующее изменение гнездового поведения. Первопричиной же является давление естественного отбора, вызванное изменением окружающей среды: исчезновение дубов и их замена соснами дают мутантному аллелю Ь преимущество перед обычным прежде аллелем а. Именно естественный отбор — причина того, что во всей популяции аллель а заменяется на аллель Ь.
Перепугать непосредственные причины с первопричинами очень легко, особенно когда дело касается такого сложного многоуровневого процесса, как эволюция человека. Мы часто читаем, что причиной революционного увеличения человеческого интеллекта было подчинение огня, или прямохождение, или переход к охоте с преследованием дичи, или сочетание каких-то из этих факторов. Я не оспариваю огромную важность этих эволюционных вех, однако первопричинами они не были. Это были этапы пути к сложному общественному поведению современного человека. Каждый из них был независимой адаптацией и имел свои собственные непосредственные причины и первопричины, подобно постоянным гнездам и постепенному провиантированию, которые подвели к эусоциальности некоторых насекомых. Заключительным шагом было формирование мозга современного Homo sapiens. Оно вызвало культурный взрыв, который продолжается по сей день.
18 | Движущие силы общественной
эволюции |
Говоря об эволюции общественного поведения, следует в первую очередь подумать о том, на каком уровне биологической организации работает естественный отбор? Может быть, он воздействует на особей таким образом, что их потомки собираются в группы и действуют сообща, — и все потому, что принадлежать к такой группе — огромное преимущество? Или, быть может, альтруизм основан на родстве, ведь член группы, способствующий размножению родственников, передает в следующее поколение свои гены, даже не оставив потомства? Или же, наконец, дело в том, что носители генов альтруизма проявляют столь высокую степень сотрудничества и организованности, что их группы неизменно одерживают верх над объединениями эгоистичных индивидов?
Результаты многих новейших исследований указывают на третий из предложенных вариантов ответа, иными словами, на то, что естественный отбор работает на групповом уровне. В этой главе я объясню, почему это так. Как и в главе 16, мне придется прибегнуть к научному стилю изложения материала, но я постараюсь, чтобы мои объяснения были максимально понятны широкому кругу читателей. Так случилось, что я много лет занимался этой темой, и один фрагмент общей теории, над которым я работал последнее время, стал предметом яростной полемики. Так что можете считать эту главу свежей сводкой с научной передовой.
Долгое время эволюцию сложного общественного поведения объясняли теорией совокупной приспособленности (она же теория родственного отбора). Согласно этой теории, центральное место в происхождении общественного поведения занимает родство. По сути она утверждает, что чем теснее родственные связи в группе, тем большего альтруизма и сотрудничества можно ожидать от ее членов и, следовательно, тем больше шансов, что группа перейдет к общественному образу жизни. Эта гипотеза обладает необычайной притягательностью, поскольку интуитивно приемлема. Это ведь так естественно — отдавать предпочтение своей родне, так почему бы родственникам — и муравьям, и людям — не держаться вместе?
На протяжении более чем сорока лет теория совокупной приспособленности оказывала глубокое воздействие на интерпретацию генетической эволюции всех форм общественного поведения. Особенно часто к ней обращались для объяснения родственного альтруизма, то есть самопожертвования (в том числе в виде отказа от собственного вклада в размножение) ради другого члена группы.
Совокупная приспособленность — продукт родственного отбора, за счет которого индивид может влиять на успех размножения своих родственников по боковой линии, например родных и двоюродных братьев и сестер. В строго биологическом смысле поведение индивида можно охарактеризовать как альтруистическое, если с точки зрения генетической приспособленности его родственники по боковой линии оказываются в выигрыше, а сам альтруист — в проигрыше. «Совокупная приспособленность» индивида складывается из его собственной приспособленности (то есть числа его потомков, которые успешно размножились и имеют собственных детей) и влияния его действий на приспособленность родственников по боковой линии (братьев и сестер, тетушек и дядюшек, двоюродных братьев и сестер). Если совокупная приспособленность индивида и приспособленность его группы в целом повышаются (несмотря на снижение
приспособленности отдельных особей), то, говорит нам теория, повышается также и частота гена альтруизма у всего вида. Теория родственного отбора быстро привлекла внимание как ученых, так и общественности, которые оценили ее простоту и убедительность, с которой она, как казалось тогда, объясняла важность альтруизма в общественной жизни.
Идею родственного отбора впервые высказал британский биолог Дж. Б. С. Холдейн в 1955 году, но сама теория была разработана несколько позже, в 1964 году, его соотечественником Уильямом Гамильтоном. Он вывел основополагающую формулу, ставшую социобиологическим аналогом «e = mc2», — неравенство гЬ > с. Оно означает, что частота аллеля альтруизма в популяции повысится, если выгода (b — benefit) для родича, умноженная на степень его родства (г — relatedness) с альтруистом, больше, чем ущерб (с — cost) для альтруиста. Холдейн и Гамильтон понимали г как долю общих, идентичных по происхождению генов у альтруиста и «реципиента» альтруизма. Например, альтруизм возникнет, если выгода для брата или сестры в два раза превышает ущерб для альтруиста (г = 1/2) или если выгода для кузена или кузины превышает ущерб в 8 раз (г = i/8). Очень грубый пример: вы способствуете распространению вашего альтруистичного гена, если из альтруистических соображений отказываетесь обзаводиться потомством, но в результате ваших действий у вашей сестры рождается втрое (вчетверо, впятеро и т. д.) больше детей.
Идею родственного отбора лучше всего выразил сам Холдейн:
«Предположим, что вы — носитель редкого гена. Он определенным образом влияет на поведение, заставляя вас броситься в реку, чтобы спасти тонущего ребенка, однако один шанс из десяти, что вы при этом погибнете. У меня такого гена нет. Я спокойно стою на берегу, и ребенок тонет на моих глазах. Если это ваш сын или дочь или же ваш брат или сестра, то вероятность того, что спасенное чадо тоже имеет этот редкий ген, равна 50%. Таким образом, на один