Химерное растение № 77/2 по числу хромосом в корешках не показало отличий от родителя — томата. Это подтверждает, что его внутренний компонент принадлежал томату.
Растение № 73/3 имело в корешках 48 хромосом (рис. 90), но один корешок имел около 70 хромосом. Фиксация корешков данного растения производилась два раза: 12 августа и 10 октября. В соматических пластинках, зафиксированных в первый раз, хромосомы производили впечатление чисто томатных, во вторую фиксацию хромосомы заметно укоротились и как бы уплотнились.
Чисто паслёновые побеги из каллюса прививок № 77 и 146 имели при летней (первой) фиксации метафазные пластинки типа паслёна. Другая картина наблюдалась при фиксации в холодные дни (вторая фиксация). В этом случае нормальных пластинок, содержащих 72 интенсивно окрашенные хромосомы, было меньшинство, в большей части пластинок присутствовали и окрашенные и бесцветные хромосомы. Наконец, так же, как и в чистом паслёне, в некоторых клетках наблюдались только хорошо окрашенные хромосомы, но в меньшем против нормы числе.
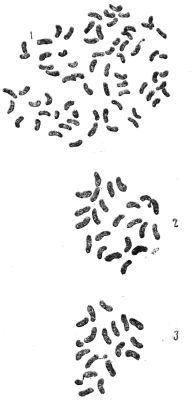
Рис. 89. Соматические хромосомы: 1 — S. nigrum — контроль, 72 хромосомы; 2 — томатный побег из каллюса прививки 77/1, 23 хромосомы; 3 — томатный побег из каллюса прививки 77/1, 17 хромосом.

Рис. 90. Соматические хромосомы растения 77/3 (48 укороченных и утолщённых хромосом).
Ещё реже нормальные паслёновые пластинки удавалось наблюдать при осенних фиксациях в паслёновых побегах, выщепившихся из изменённых (химерных) растений[8]. Цитологически были исследованы два таких выщепенца из растений № 77/2 (рис. 91) и 146 (рис. 92). Один из них дал около 15 % пластинок с 72 нормально красящимися хромосомами, другой — всего 1, 67 %. Основная масса исследованных пластинок у этих выщепенцев состояла из клеток, где часть хромосом оставалась слабо окрашенной или совсем неокрашенной. Ниже приводится таблица 77, содержащая данные сравнительного анализа метафазных пластинок чистого паслёна, побега из каллюса и двух паслёновых побегов из изменённых растений. Если в чистом паслёне нормальных пластинок с 72 хромосомами было 73,77 %, то в паслёновом побеге из каллюса их было 30 %, а в паслёновых выщепенцах число их падает до 15,15 и 1,67 %. В полученных из каллюса паслёновых, томатных и промежуточных (химерных) побегах исследовалось также редукционное деление.
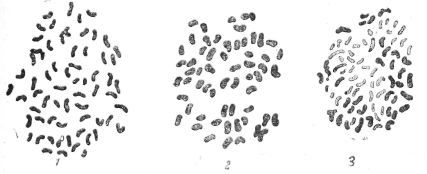
Рис. 91. Метафазные пластинки в корешках выщепенцев из растения 77/2 (прививка Solarium nigrum на Жёлтый Мичурина): 1 — 63 хромосомы; 2 — 60 утолщённых, укороченных хромосом; 3 — 34 окрашенные хромосомы, остальные неокрашенные или слабо окрашенные.
Во всех растениях типа паслёна и типа томата, наряду с нормальным редукционным делением, наблюдались многочисленные картины, свидетельствующие о расстройстве его механизма. Резче всего последствия ненормально прошедших мейотических делений видны на стадии тетрад. Здесь встречаются материнские клетки с тремя вместо четырёх ядер, с ядрами неодинаковой величины, с одним или несколькими микроядрами, с одним гигантским ядром и т. д. Дефективные тетрадные клетки не разъединяются при лёгком нажатии на покровное стекло, как это бывает при нормальных тетрадах, а остаются соединёнными, составляя как бы плотную ткань. Это напоминает молодую археспориальную ткань до начала редукционного деления с тем различием, что клетки дефективных тетрад содержат лишь по небольшому комочку съёжившейся плазмы. Весь вид этих тетрадных клеток представляет картину явной дегенерации. Некоторые клетки, кроме четырёх ядер, содержат отдельные хромосомы, оставшиеся не включёнными в ядра и затерявшиеся в плазме. Широко распространено на этой стадии также слияние ядер, о котором говорит ряд переходных картин. Вначале два ядра близко соприкасаются, затем на месте соприкосновения их ядерная оболочка растворяется. Общее крупное ядро имеет овальную или почковидную форму и два ядрышка, которые после сливаются в одно, и ядро постепенно приобретает нормальную округлую форму. Многие пыльники и целые цветки содержат сплошь дегенеративную пустую пыльцу со смятыми сморщенными оболочками. В других цветках наблюдалось и более нормальное редукционное деление с экваториальными пластинками, содержащими нормальное число хромосом, и с нормальными тетрадами и пыльцой.
Эта разная степень мейотической неустойчивости в отдельных цветках безусловно связана с тем, что растения, в результате прививки выйдя из состояния относительного равновесия, становятся особенно чувствительными ко всем изменениям внешней среды и реагируют на них более резко, чем растения, находящиеся в устойчивом состоянии. Условия питания, складывающиеся по-разному в различных цветках одного и того же растения, могут вызвать разную степень расстройства мейозиса в организмах с нарушенной инерцией наследственности.
Характерной чертой редукционного деления в клетках подопытных растений является независимое друг от друга поведение отдельных материнских клеток пыльцы, выражающееся в нарушении синхронности при прохождении стадий деления, что, очевидно, должно обусловить и большее разнообразие будущих гамет, а следовательно, и большее разнообразие семенного потомства.

Рис. 92. Растение № 146 (прививка S. nigrum на Альбино черенок, давший промежуточный плод). Клетка имеет 68 окрашенных и 4 слабо-окрашенных хромосомы.
Во всех случаях, когда редукционное деление идёт с большими нарушениями, резко изменяется отношение материнских клеток к употребляемым фиксаторам и окраскам. Обработка уксуснокислым кармином, дающая у контрольных растений прекрасную диференциальную окраску ядра и плазмы, в описываемых случаях резкого расстройства редукционного деления давала сплошное густо розовое окрашивание, зачастую не позволяющее даже определить стадию деления. Вероятно, отдалённые прививки сопровождаются глубоким изменением химизма клетки, что проявляется совершенно иным отношением её к фиксирующим и окрашивающим реактивам.
Характерной особенностью молодых пыльцевых зёрен у растений, полученных из каллюса отдалённой прививки, является также то, что они долгое время спустя после расшнурования плазмы на четыре клетки остаются соединенными в четверки, так что пыльца имеет вид тетрад, хотя каждое пыльцевое зерно уже покрыто собственной оболочкой и имеет нормальную величину. Все эти черты резко отличают характер редукционного деления у растений, выросших из каллюса, от обоих исходных чистых видов.
В соответствии с описанными проявлениями расстройства механизма мейозиса находится и качество зрелой пыльцы. Лучшим показателем зародышевой или мейотической неустойчивости служит процентное соотношение нормальных и абортивных пыльцевых зёрен. Вычислялось оно на основании данных подсчёта тех и других в нескольких полях зрения микроскопа, взятых из разных мест препарата. Число сосчитанных таким образом пыльцевых зёрен было 200–300. В таблице 78 показан процент хорошей пыльцы у отдельных растений, выращенных из прививочного каллюса.
Приведённые данные характеризуют не растение в целом, а только отдельные цветки, между которыми (на одном и том же растении) по проценту жизнеспособной пыльцы могут существовать большие колебания.
Ту же картину аномалий в прохождении редукционного деления материнских клеток на разных этапах его представляют побеги чистых видов (паслёна и томата), выщепившихся на изменённых растениях. Общее количество пыльцы у выщепенцев по сравнению с побегами из каллюса уменьшено. Очевидно, большое количество пыльцевых зёрен отмирает в период первого и второго редукционных делений. Процент абортивной пыльцы по отдельным цветкам сильно варьирует (в большинстве цветков он колеблется от 35 до 60), но есть пыльники почти полностью с нормальной пыльцой и пыльники и целые цветки, сплошь занятые дегенерирующей деформированной пыльцой или уже одними смятыми пустыми оболочками.
8
На факты разной окрашиваемости хромосом в зависимости от условии среды указывают и другие авторы (1940; Wilson, 1942).