По многочисленным данным, штаммы FI- легко селекционировать из популяции FI+ при помощи специфической антисыворотки. Такие штаммы не образуют видимой капсулы, не агглютинируются капсульной антисывороткой, не высвобождают FI после обработки их ультразвуком и не вызывают образования соответствующих антител.
Помимо явных FI+ и FI- штаммов встречается еще третий тип штаммов, обладающих свойствами как первого, так и второго типа (штаммы FI(). Штаммы FI(не способны к образованию видимой капсулы ни на питательных средах, ни в организме животных. Однако такие штаммы способности к синтезу FI полностью не лишены, о чем можно судить с помощью реакции преципитации в геле или по индукции ими специфических антител. Два таких штамма были получены путем селекции, а один выделен от человека, умершего от чумы (штамм Bryan [Burrows T. W., Bacon, G. A., 1958].
По химической природе FI оказался белковым агрегатом с мол. массой 300 кДа, состоящим из двух компонетов с одинаковыми антигенными свойствами. Один из них, изоэлектрическая точка которого лежит при рН 4,6, соединен с олигомерным галактаном, т. е представляет собой гликопротеин, тогда как второй, с pI 4,8, является чистым белком. Оба компонента распадаются на субъединицы с мол. массами от 15 до 17 кДа [Bennet L., Tornabene T., 1974] и легко образуют исходный, высокомолекулярный комплекс. Упаковка молекулы FI происходит за счет водородных и гидрофобных взаимодействий без образования дисульфидных связей [Наумов А. В., Самойлова Л. В., 1995]. B-клеточный эпитоп, доступный для антител, выглядит как гидрофильная петля на поверхности полимерной молекулы [Zav'yalov V. et al., 1995a].
Что касается генетического контроля синтеза FI, то, по данным ряда авторов, например О. А. Проценко и соавт. [1983], он осуществляется «крупной» плазмидой (pFra), молекулярная масса которой колеблется от 60 до 65 мДа. Фрагмент pFra Y pestis EV, непосредственно связанный с синтезом FI, (оперон fI (недавно был детально изучен. Он оказался относительно небольшим (около 4 мДа) и включает в себя 4 гена (рис. 11). Клетки кишечной палочки, несущие рекомбинантную плазмиду с цельным опероном fI, образуют капсулу, содержащую FI [Karlyshev A. et al., 1996]. Примечательно, что эти клетки обладают способностью связывать интерлейкин 1β(hiL-1β), причём ответственным за связывание является белок CaFIA; этот факт должен учитываться при анализе роли капсульного вещества в патогенезе чумы [Zav'yalov V. et al., 1995b].
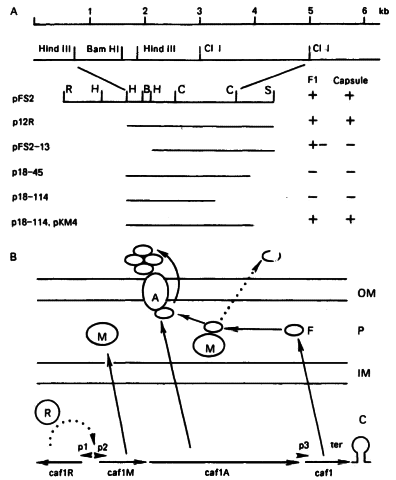
Рис. 11. Физическая и генетическая карта кластера генов биосинтеза капсулы Y. pestis [Karlyshev А. Е. et al., 1996].
А — структура различных делеционных вариантов плазмиды pFS2 и фенотипы штаммов Е. соіі, несущих соответствующие плазмиды: R — EcoRI, S — SalGI, В — BarnHI, С — СП, Н — Hindlll; В — генетическая организация и предполагаемая регуляция экспрессии кластера генов и схематическое изображение роли продуктов соответствующих генов в секреции субъединиц и образовании капсулы. Продукты генов: cafIM (М), caflA (A), cafl (F), cafIR (R). ОМ — внешняя мембрана, Р — периплазматическое пространство, ІМ — внутренняя мембрана, С — цитоплазма.
Казалось бы, все ясно. Однако у штамма Yreka в образовании FI может участвовать плазмида с мол. массой 13 мДа, отсутствующая у других штаммов чумного микроба [Tsukano H et al.,1986]. Чтобы убедиться в этом, японские авторы прибегли к помощи комбинации элиминирующих плазмиды агентов и получили клетки штамма Yreka, которые наряду с указанной и еще четырьмя плазмидами утратили поверхностный капсульный материал [Tsukano H., 1989]. Но полученные клетки оказались неоднородными; одни были лишены FI, a другие напоминали штаммы FI(и содержали FI, но только в цитоплазме. Поскольку оба типа клеток были бесплазмидными, авторы пришли к выводу, что именно плазмида с мол. массой 13 MД связана с появлением поверхностного капсульного антигена и не имеет отношения с синтезу внутриклеточного антигена. Дополнительные опыты дали указание на наличие каких-то других генетических элементов, кодирующих образование внутриклеточной FI.
Вопрос о генетическом контроле синтеза FI окончательно решеным считать нельзя. Помимо сказанного и собственных неопубликованных данных, в этом нас убеждают также результаты исследований других авторов, в частности, M. S. Zhao и соавт. [1990]. По мнению последних, соответствующие гены располагаются на транспозоне. Иначе трудно объяснить тот факт, что у одних штаммов чумного микроба гены FI связаны с крупной плазмидой, а у других (с хромосомой. И еще один вопрос, на который предстоит ответить: если синтез FI действительно связан со столь небольшим фрагментом pFra, как утверждают A Karlyshev и соавт., то что же кодирует остальная часть этой неконъюгативной плазмиды? Мышиный токсин, о чём пишут другие авторы? Как нам кажется, ответы на поставленные вопросы принципиально важны, поскольку они помогут понять причину, породившую сомнения в FI, как в одном из факторов вирулентности чумного микроба [Davis K. J. et al., 1996].
pH6-антиген (pH6Ag) был описан S. Ben-Efraim и соавт. [1961]. Они показали, что этот антиген образуется чумным микробом in vitro и in vivo при 37 °C и рН ниже 6,7, т. е. при условиях которые сходны с таковыми в фаголизосомах и абсцессах. По данным L. Bichowsky-Slomnicki и S. Ben-Efraim [1963], этот антиген сообщает клеткам чумного микроба большую стабильность в суспензиях, агглютинирует эритроциты и обладает цитотоксичностью, что может иметь непосредственное отношение pH6Ag к вирулентности, поскольку мутации в хромосомном структурном (psaA)или регуляторном (psaE) гене приводят более чем к 100-кратному увеличению ЛД50 [Linder L. E. et al., 1990].
pH6Ag образует фимбриоподобные поверхностные структуры, построенные из субединиц с мол. массой 15 кДа. Их образование индуцируется внутри макрофагов. Небезынтересно, что в отличие от гена psaA, «работа» гена psaE не зависит от температуры и pH среды. Одной из особенностей этих структур (белка PsaA является их способность играть роль Fc-рецепторов; они связываются с нормальными IgG человека, что приводит к возникновению псевдоиммунных комплексов, и, как следствие, к антигенной мимикрии [Zav'yalov V. et al, 1996]. Возможно, что утрата вирулентности в опытах L. E. Linder и соавт. [1990] обусловливалась именно тем, что их мутанты не могли «обманывать» иммунную систему организма, как это делают неизмененные штаммы.
С помощью ДНК гибридизации и иммуноблотинга антиген, подобный pH6-антигену, выявлен у возбудителя псевдотуберкулёза [Linder L. E., Tall B. D., 1993], у которого он также связан с вирулентностью [Muhr J., 1993; цит. по Holmstr(m A., 1995]. Кроме того, 44 %-гомология с pH6-антигеном была обнаружена у Myf-белка (мукоидного фактора) Y. enterocolitica [Iriate M. et al., 1993]. В то же время, хромосомные локусы, кодирующие pH6-антиген и Myf-белок, имеют сходство с локусами Pap-пилей кишечной палочки (papC papD, ответственными за транспорт и сборку субъединиц пилина, что интересно с эволюционной точки зрения.
При изучении потребностей в источниках питания авирулентного штамма FS и его вирулентного мутанта MP6 было замечено [Jackson S., Burrows T. W., 1956], что оба штамма на синтетической среде с гемином образуют пигментированные колонии. После 4-дневной инкубации от пигментированных колоний (Р+) отщепляются вторичные непигментированные колонии (Р-), которые остаются непигментированными и в последующих генерациях (превращение штаммов Р- в Р+ никогда не происходит).
Как установил [Burrows T. W., 1960 a, b], превращение штамма Р+ в штамм Р- сопровождается потерей вирулентности для мышей, даже если штамм Р(сохраняет способность к образованию других детерминантов вирулентности.
Потеря вирулентности, связанная с утратой способности образовывать пигментированные колонии на среде с гемином, оказалась обратимой и полностью восстанавливалась, если штамм P- (c другими детерминантами вирулентности) вводили мышам вместе с нетоксичными дозами гемина или солей железа. В опытах на морских свинках этот эффект проявлялся слабее. Заменить соли железа солями других металлов не удалось. Железо не возвращало вирулентность пигментированным авирулентным штаммам, например, P+ FI+ VW-, и их непигментированным мутанатам P- FI+ VW- [Jackson S., Burrows T. W., 1956].