В настоящее время широко применяется метод отведения электрических потенциалов от коры головного мозга. При помощи специальных усилителей удается записать на бумаге или фотопленке электрические токи, возникающие в различных отделах головного мозга. Нормальная активность мозга характеризуется определенным ритмом электрических колебаний и закономерной их величиной. При сильных болевых раздражениях электрическая активность коры мозга ослабевает, величина ритмичных колебаний (так называемых альфа-волн) уменьшается. Анализ полученных данных показывает, что при боли усиливаются процессы возбуждения по всей коре мозга и в то же время в ней появляются отдельные очаги застойного возбуждения. Эти изменения наиболее выражены по нисходящему ряду в теменных, затылочных, височных и лобных долях мозга. Однако они не специфичны для боли. Аналогичная картина была отмечена при сильных температурных воздействиях и даже при поглаживании кожи, прикосновении к ней и т. д.
Помимо коры головного мозга, огромное значение для восприятия болевого ощущения имеют зрительные бугры.
«В физиологии нервной системы, — писал В. М. Бехтерев в 1883 г., — нет более темной области, как отправление зрительных бугров». Несмотря на то, что за истекшие годы появилось огромное количество работ, посвященных анатомии и физиологии зрительных бугров, роль этой области мозга далеко еще нельзя считать окончательно выясненной. Лишь в последние годы некоторые стороны деятельности зрительных бугров стали значительно яснее и стала более понятной их роль в физиологии больших полушарий мозга.
Зрительные бугры представляют собой большие скопления серого вещества в межуточном мозгу. С двух сторон, с правой и левой, они образуют стенки третьего желудочка мозга с огромным количеством самых разнообразных нервных путей, которые связаны почти со всеми отделами головного, продолговатого и спинного мозга.
На разрезе легко обнаружить, что в зрительных буграх имеется пять основных серых ядер, разделенных прослойками белого вещества. К передней части внутреннего (медиального) ядра подходят волокна обонятельного тракта, к задней его части — волокна протопатической (см. стр. 132) болевой чувствительности. Здесь по современным представлениям формируются чувства «удовольствия» и «неудовольствия». Задняя часть наружного (латерального) ядра является основным центром проприоцептивной чувствительности, т. е. получает импульсы из мышц и суставов, а также тактильные, болевые и температурные сигналы.
Остальные ядра не имеют непосредственного отношения к восприятию боли. Роль их многообразна и зависит в значительной «степени от связей с другими отделами мозга.
Бесконечное число нервных волокон соединяет ядра зрительных бугров с ядрами подбугорья, где заложены центры высших вегетативных центров (обмена веществ, терморегуляции, сосудистого тонуса и т. д.) с другими подкорковыми образованиями и, в первую очередь, с корой головного мозга. Чтобы убедиться в этом, достаточно взглянуть на рисунок 14.
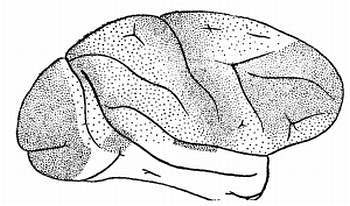
Рис. 14. Окончания бугрово-корковых путей в мозгу обезьяны макаки
Каждая точка, изображенная на поверхности коры головного мозга обезьяны макаки, показывает окончание пучка волокон, соединяющих зрительные бугры с корой. Эти волокна пронизывают всю толщу мозга. Они обеспечивают бесперебойную двустороннюю связь от зрительных бугров с корой и передают импульсы в обе стороны: от клеток коры к зрительным буграм и обратно.
В течение многих лет дебатировался вопрос о расположении высших центров болевого ощущения. До недавнего времени считалось наиболее вероятным, что болевое ощущение формируется не в коре головного мозга, а в зрительных буграх. Это предположение поддерживали многие виднейшие физиологи и невропатологи мира. Впервые его высказали английские ученые Гэд и Холмс, а в дальнейшем французы Лавастин и Лериш, русские исследователи Аствацатуров и Орбели, а за ними и многие другие. В то же время большинство исследователей склоняется к мысли, что эта область мозга является лишь важнейшим, узловым, но в то же время промежуточным центром болевого восприятия.
Зрительные бугры представляют древнейшую часть головного мозга. Они образовались в нервной системе живых существ, населявших землю за много миллионов лет до появления человека. Лишь в процессе эволюции, на более высоких ступенях зоологической лестницы появилась та часть мозга, которую мы называем полушариями.
Спор между сторонниками «корковой» и «бугровой» локализации болевого восприятия потерял в настоящее время свою остроту. Вскрылись новые обстоятельства, позволившие по-новому проследить путь болевых ощущений, совершенно иначе расценить давно известные факты и установившиеся, как казалось многим, «ученические» истины.
Уже давно известно, что при раздражении какого-нибудь чувствующего нерва в коре головного мозга можно обнаружить два вида электрической активности, различающихся временем их возникновения.
В 1942 г. американские ученые Морисон и Демпси показали, что и при электрическом раздражении ядер зрительного бугра в коре можно получить два типа электрических реакций. Первый с коротким скрытым периодом, обнаруживающимся преимущественно в чувствительных зонах коры, в корковых ядрах тех или иных анализаторов. Второй — с длинным скрытым периодом, распространяющимся по всей коре и не поддающимся точной локализации. Эта распространенная (генерализованная) реакция коры головного мозга наблюдается при раздражении всех видов чувствующих волокон — зрительных, слуховых, обонятельных, тактильных, температурных и болевых. Можно предположить, что периферические раздражения поступают в кору по двум различным проводящим системам. Один путь — специфический, знакомый нам во всех деталях. Другой — до недавнего времени гипотетический, мало изученный.
Используя специальные так называемые стереотаксические дриборы, физиологи научились вводить тончайшие, невидимые невооруженным глазом электроды не только в скопления нервных клеток, расположенные в глубинных отделах мозга, но даже в отдельные нейроны.
Электрический ток, пропущенный через такие электроды, способен раздражать нервные ядра мозга, а в случае необходимости разрушать строго ограниченные участки мозговой ткани. При этом экспериментатор имеет возможность наблюдать, каким становится поведение животного, как изменяются в его организме физиологические и биохимические процессы.
Хирурги во время операции на центральной нервной системе давно заметили одно интересное обстоятельство.
Оказалось, что можно во время операции производить любые разрезы через кору больших полушарий мозга, можно удалять значительные участки мозговой ткани, иногда даже целое полушарие, тем не менее пациент продолжает бодрствовать и не теряет сознания. Однако стоит лишь слегка задеть инструментом некоторые точечные участки в глубине мозга, как больной немедленно впадает в бессознательное состояние, похожее на сон. В конце 40-х годов нашего столетия два видных физиолога — американец Г. Мэгоун и итальянец Д. Моруцци — провели на животных серию экспериментов, которые не только объяснили это явление, но и заложили основу изучения функций особого образования мозга, известного под названием ретикулярной формации.
Ретикулярная формация представляет скопление нервных клеток в центральных отделах той части головного мозга, которую называют мозговым стволом. Нервные клетки ретикулярной формации, благодаря большому числу ветвящихся и переплетающихся отростков, образуют густую нервную сеть, откуда и возникло название «ретикулярная» или в переводе на русский язык сетевидная формация. Анатомы уже давно описали этот своеобразно построенный отдел мозга. Но значение его стало понятным лишь сравнительно недавно.