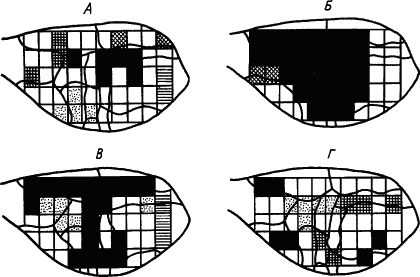
Рис. 15. Динамика синхронизации волн электрокортикограммы кролика в процессе выработки условного рефлекса. А — исходный фон; Б — после 12 сочетаний; В — после 60 сочетаний (появился условный рефлекс); Г — после 90 сочетаний (прочный условный рефлекс):
квадратами обозначены участки отведения; одинаково изображены участки, синхронность волн между которыми сохраняется не менее 70 % времени измерения
Многие сведения, существенные для понимания механизма образования временных связей, были получены при исследовании разных режимов условного раздражения и подкрепления, а также пространственных характеристик регистрируемой активности. Так, успех образования временной связи по показателям синхронизации усвоенных ритмов оказался зависимым главным образом от четырех факторов: 1) изоритмии сигнала и подкрепления; 2) оптимальности их частоты (для кролика 4–7 Гц); 3) отношения фаз (синфазность) и 4) достаточного объема нервного субстрата, вовлеченного в этот процесс. Отсюда сделано заключение, что наблюдаемая при выработке условных рефлексов синхронизация электрических потенциалов в разных структурах мозга отражает перестройку их лабильности, которая делает возможным движение нервных процессов между ними. Временная связь формируется на основе такого «проторения» пути синхронизированными ритмами биоэлектрической активности (М.Н. Ливанов, 1980).
Значение функционального состояния лабильности и возбудимости структур, участвующих в образовании временной связи, показано в опытах с созданием искусственных очагов повышенной возбудимости, имитирующих явления доминанты — усиленного ответа на раздражитель по сравнению с другими областями. В таких очагах быстрее и легче усваиваются и могут долго сохраняться сигнальные ритмы, распространяющиеся на другие активируемые структуры.
Доминанту рассматривают в качестве основы для формирования временных связей, как переходную форму от суммационного к настоящему условному рефлексу. В доминантном очаге возникают сложные взаимоотношения между нервными и глиальными клетками. Формирование доминанты имеет динамичный характер. Она проходит ряд стадий, среди которых стадия специализации имеет особое значение для организации целенаправленного поведения. О функциональном состоянии мозга, влияющем на течение условно-рефлекторных реакций, определенное представление дает состояние основной ритмики ЭКоГ, на фоне которой подается сигнал.
Образование условных рефлексов, как уже упоминалось, находит отражение в электрической активности не только коры, но и подкорковых структур. При одновременной регистрации потенциалов корковых областей, ядер таламуса и ретикулярной формации среднего мозга в процессе образования условного рефлекса отмечено, что условное возникновение реакции десинхронизации или усвоение ритма сигнального раздражителя может появляться в подкорковых структурах, в частности в ретикулярной формации, раньше, чем в коре (рис. 16), а разрушение неспецифических ядер таламуса устраняет возникшую условную реакцию.
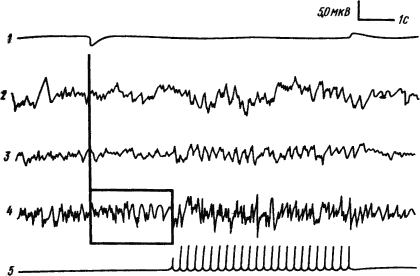
Рис. 16. Условно-рефлекторное воспроизведение ритма сигнального раздражения в ретикулярной формации среднего мозга кошки (отмечено квадратом) (по Н. Иошии и др.):
1 — отметка сигнального звукового раздражителя, 2 — переднее вентральное ядро таламуса, 3 — срединный центр, 4 — ретикулярная формация, 5 — подкрепляющий ритмический световой стимул
На этом основании было высказано предположение, что временные связи условного рефлекса образуются на подкорковом уровне в неспецифических структурах мозгового ствола. Полагали, что при сочетании условного и безусловного раздражителей по коллатералям их специфических путей происходит конвергенция возбуждений в неспецифические структуры ретикулярной формации среднего мозга. Так объясняли образование их связей, а ослабление восходящих активирующих влияний считали причиной ограничения области десинхронизации в коре. Однако наблюдения за динамикой электрической активности корковых и подкорковых структур в процессе выработки и укрепления условного рефлекса показали, что активность неспецифических образований мозгового ствола наиболее выражена в начальных стадиях выработки, когда они вызывают диффузное возбуждение, а по мере упрочения рефлекса на первый план выступают специфические системы на уровне коры (показано в опытах на кошках и собаках).
Деятельность неспецифических структур мозга, в частности восходящей активирующей системы ретикулярной формации среднего мозга, как показали исследования, действительно играет важную роль в образовании временных связей, обеспечивая высокую степень работоспособности высших отделов мозга. Замыкание временных связей является многоуровневым процессом, в который вовлекаются как кора, так и подкорковые образования, причем большую роль в этих процессах играет гиппокамп.
Широкое участие различных структур мозга в образовании временных связей проявляется и в синхронизации ритмов электрической активности корковых областей и многих подкорковых структур. Однако функция большинства из них, особенно неспецифических областей, связана с созданием условий для образования временных связей. Это достигается активацией высших отделов мозга и распространением этого активного состояния на обширные области коры. По мере укрепления связей эти функции отпадают и участие неспецифических областей становится все более ограниченным. Многие электрические показатели этих процессов, резко выраженные в начале выработки рефлекса, в дальнейшем ослабляются и могут исчезнуть. Так происходит с условной реакцией десинхронизации ЭЭГ человека при упрочении условного двигательного рефлекса. Как видно из рис. 17, четкая депрессия альфа-ритма в отведении, соответствующем корковому участку движения руки, по мере увеличения числа сочетаний становится все меньше и при прочном рефлексе совершенно исчезает.
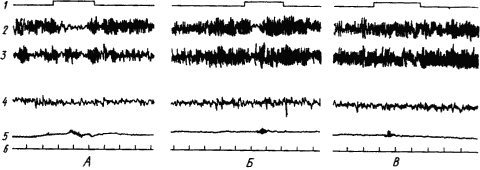
Рис. 17. Изменение ЭЭГ человека при выработке условного двигательного рефлекса на звук метронома. А — после 10 сочетаний; Б — после 28 сочетаний; В — после 80 сочетаний (по А.Б. Когану):
1 — включение метронома, 2, 3 — отведение от левой и правой роландовых борозд, 4 — от височной коры, 5 — электромиограмма общего разгибателя пальцев руки, реагирующей движением, 6 — отметка времени (1 с)
Исследование фоновой ритмики при выработке условных пищевых и оборонительных рефлексов у кроликов в условиях свободного поведения показало учащение колебаний ЭКоГ и укорочение периода условного ответа. При этом наблюдали сходный характер изменений электрической активности при пищевом и оборонительном рефлексах. В специальных опытах с созданием пищевой (голодание) или оборонительной (болевое раздражение) доминанты в коре, ретикулярной формации и лимбических структурах возникали синхронизированные колебания, отражающие высокий уровень неспецифической активации. Эта активация проявлялась не только в учащении импульсов ответов нейрона на условные раздражители, но и в периоды между их предъявлением (рис. 18). Так в деятельности нейронов отражались фазические и тонические компоненты ее перестроек при образовании как пищевых, так и оборонительных условных рефлексов. Однако прямо противоположная реакция в случае пищевого или болевого подкрепления сигнала означает, что проявившаяся в электрической активности неспецифическая активация избирательно действует на структуры мозга, реализующие процессы разного биологического значения. Пищевое подкрепление условного рефлекса придает ему эмоционально-положительный, а оборонительное — эмоционально-отрицательный характер.