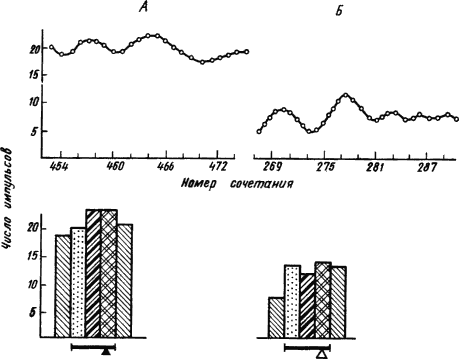
Рис. 18. Тонические и фазические импульсные реакции нейронов гиппокампа при условном пищевом (А) и оборонительном (Б) рефлексах (по Б.И. Котляру, Н.О. Тимофеевой):
А — уровень фоновой активности между сочетаниями, Б — усредненное количество импульсов за 1 с в разные этапы условно-рефлекторной деятельности; чертой отмечено время действия условного раздражителя, треугольником — момент условной реакции
Процессы образования временных связей находят отражение и в сверхмедленных колебаниях потенциалов (СМКП). Так, при выработке у кролика условного оборонительного рефлекса на световой сигнал, сопровождаемый электрокожным подкреплением, наблюдали закономерные изменения СМКП. При этом в начальный период увеличиваются амплитуда и частота СМКП в коре и ряде подкорковых структур, достигая максимума при генерализации рефлекса. Прочное его закрепление сопровождается уменьшением СМКП до полного исчезновения, а угашение рефлекса вновь их вызывает. При выработке условных рефлексов у кроликов происходил сдвиг потенциалов коры вначале в сторону позитивности, а затем негативности, нарастающей по мере укрепления рефлекса. Такую негативизацию рассматривают как показатель деполяризации, обусловливающей местное возбуждение.
Обнаружение связи медленных электрических процессов с психической деятельностью человека (Н.П. Бехтерева, 1966) было исходным для целого направления исследований их динамики, например при обучении иностранным словам или использовании для анализа нейрофизиологических механизмов мнестической деятельности и процесса формирования матрицы долгосрочной памяти. В последнее время СМКП связывают с метаболическими процессами, влияющими на электродный потенциал, а их сдвиги, наблюдаемые при выработке условного рефлекса, обозначают как колебания биоэлектрохимической активности.
Таким образом, при формировании временных связей динамика электрических реакций десинхронизации основной ритмики сверхмедленных потенциалов и усвоения ритма условного раздражителя отражает вначале широкий охват возбуждением неспецифических и специфических систем на корковом и подкорковом уровнях. Он соответствует стадии генерализации условного рефлекса, по мере закрепления которого происходит сосредоточение этих электрических реакций в специфических структурах корковых проекций сигнала и двигательного ответа, а также в подкорковых образованиях.
Вызванные потенциалы. Рассмотренное выше явление усвоения ритмов раздражения представляет собой серии укороченных вызванных потенциалов, в которых полное развитие каждого из них прерывается возникновением следующего. В более полном виде вызванные потенциалы (ВП) с их первичными ответами и поздними компонентами выявляются при относительно редких (1–2/с) раздражениях и могут быть использованы как показатели динамики процессов высшей нервной деятельности. В результате многочисленных исследований изменений ВП в зоне сигнального раздражителя при образовании временных связей накопились разнообразные и во многом противоречивые сведения о закономерностях этих изменений.
Так, в первых исследованиях динамики ВП проекционных зон сигнального раздражителя при выработке условного рефлекса одни авторы отмечали его увеличение, другие — уменьшение, третьи — вообще ставили под сомнение наличие закономерной корреляции между ходом образования условного рефлекса и изменениями ВП. Видимо, причина противоречивых результатов состояла в том, что изменения ВП в чрезвычайной степени зависят от стадии образования условного рефлекса (генерализованного или специализированного, непрочного или упроченного), от локализации отведения (проекция условного или безусловного раздражения, первичные или вторичные сенсорные зоны), от вида, возраста и состояния объекта.
Дальнейшие исследования наглядно показали наличие такой зависимости. Изменения ВП происходят по стадиям формирования условного рефлекса. В большинстве случаев начало образования временной связи может сопровождаться некоторым снижением величины ВП. На стадии генерализации условного рефлекса иррадиация возбуждения в коре мозга проявляется в виде усиления и широкого распространения ВП почти на все области коры и укорочения латентных периодов их ответов на раздражители.
На стадий упрочения условного рефлекса происходит ограничение области возникновения ВП, отражающее концентрацию процессов возбуждения в его исходном очаге. При этом величина и форма ВП приближаются к исходным. Обобщение результатов исследования динамики ВП в сопоставлении с другими показателями выработки условных рефлексов позволило заключить, что процесс отражения окружающего мира меняет свой характер на разных этапах условно-рефлекторной деятельности. Например, начальное увеличение с последующим снижением негативной волны ВП соответствует восстановлению угашенной реакции внимания. Возрастание амплитуды негативной волны идет параллельно приобретению условным раздражителем сигнального значения.
Сравнение динамики ВП в разных проекционных зонах коры показало, что его уменьшению в зоне проекции сигнального раздражителя может соответствовать возрастание величины ВП в зоне проекции безусловного, служащего подкреплением. Однако в случае инструментального условного рефлекса отношения усложняются. Так, в зоне проекции конечности, выполняющей инструментальное действие, ВП концентрируются на очень ограниченном участке, но оказываются более выраженными в области коркового представительства сигнала. В начальных стадиях образования временных связей ВП указывают на преобладание активности первичных сенсорных зон, потом возрастает роль вторичных.
Особое значение за последнее время приобретает изучение динамики поздних компонентов ВП, следующих за первичным ответом, его позитивных и негативных медленных волн. Так, при выработке зрительных оборонительных условных рефлексов отмечались закономерные перестройки именно поздних волн ВП на вспышки света. Особенно выраженные изменения происходили в медленной негативной волне. Наряду с изменениями формы и амплитуды основных компонентов ВП появлялись новые, ранее не имевшие места волны потенциалов. Если сигналом для условного оборонительного рефлекса сделать низкочастотное раздражение неспецифических ядер таламуса, то в комплексе корковых потенциалов вовлечения по мере выработки рефлекса формируется дополнительная поверхностно-положительная волна, которая при его угашении постепенно исчезает. В то же время поверхностно-отрицательная волна ВП, регистрируемого в ограниченном участке коркового представительства конечности, совершающей оборонительное движение, оказывается более стабильной. Выработка оборонительного условного рефлекса на свет при подкреплении электрокожным раздражением приводит к тому, что ВП на оба эти стимула становятся сходными по своей форме.
Таким образом, динамика вызванных потенциалов, так же как и фоновой ритмики, отражает процессы иррадиации и концентрации возбуждения, главным образом в структурах коры разной глубины, и характер их связи с подкоркой. Поздние и дополнительные компоненты, возможно, связаны с процессами формирования временных связей.
Реакции нервных клеток. Применение микроэлектродов, погружаемых в нервную ткань, позволило исследовать электрическую активность отдельных нейронов при условно-рефлекторной деятельности. Для этого держатель микроэлектрода с регулятором его погружения укрепляли на голове подопытного животного, например обезьяны. Когда был выработан условный оборонительный рефлекс отдергивания руки на вспышки света, то импульсные ответы корковых нейронов на эти вспышки отличались от ответов до выработки условного рефлекса. Характерно, что изменения импульсных ответов у одних нейронов протекали в сторону усиления разряда, а у других — в сторону его ослабления. В этом проявляется сложная функциональная организация коркового процесса условного возбуждения как динамической мозаики возбужденных и заторможенных нейронов.