Известны многочисленные попытки выделить по свойствам нейронов клетки, участвующие в образовании временных связей. Такими свойствами считали высокую частоту фоновой импульсации, большую реакцию на условный раздражитель, способность реагировать на раздражители разной модальности, полисенсорность, высокую лабильность, способность к пластическим перестройкам — избирательную полифункциональность, сходство реакции на условный и безусловный раздражители, ответы преимущественно на подкрепление. На основании опытов главным образом с нервной системой моллюсков возникло представление о ведущей роли в образовании новых форм поведения так называемых командных нейронов, осуществляющих запуск координированных двигательных актов.
Что же происходит с нейронами, участвующими в замыкании временной связи? Давно замечено, что после выработки условного рефлекса возрастает и величина безусловного рефлекса, хотя сила подкрепляющего раздражителя остается прежней. Отсюда следует, что выработка условного рефлекса связана с повышением возбудимости нервных клеток, образующих новый рефлекторный путь. Повышение возбудимости корковых структур, участвующих в образовании условно-рефлекторной связи, можно обнаружить и непосредственно, измеряя пороги их раздражения.
После вживления электродов в область двигательного анализатора (например, кошки), можно измерять порог прямого раздражения и определять изменения возбудимости исследуемых нервных клеток мозга у животного при выработке условного рефлекса. Оказалось, что процесс выработки двигательного условного рефлекса выражается прежде всего стойким повышением возбудимости клеток двигательного анализатора (рис. 12). Только когда возбудимость корковых клеток повышается до известного уровня, появляются двигательные признаки условного рефлекса — сгибание лапы. Каждый очередной сигнал вызывает быстрый дополнительный подъем возбудимости нейронов двигательного анализатора. Если же отменить условный рефлекс, то возбудимость нервных клеток снижается до уровня, предшествующего началу выработки рефлекса. Соответственно изменяются импульсация и ответы на условный раздражитель. При этом меняется деятельность нейронов.
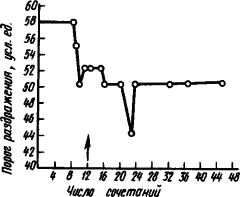
Рис. 12. Изменение порога прямого раздражения сенсомоторной коры при выработке условного рефлекса сгибания лапы у кошки (по Н.И. Николаевой):
стрелкой обозначен момент образования условного рефлекса
Так, полисенсорные нейроны, которые наиболее часто перестраиваются при образовании условного рефлекса, могут стать моносенсорными или не участвовать в условных реакциях. Исходя из того что при разных видах условно-рефлекторной деятельности в разрядах нейронов лобной коры доминируют межимпульсные интервалы каждый раз определенной длительности, было высказано предположение что положительный условный рефлекс и его торможение кодируются различными классами межимпульсных интервалов.
Электрические потенциалы, возникающие при деятельности нервной системы, могут служить тонким показателем происходящих в ней процессов. Поэтому в изучении процессов высшей нервной деятельности электрофизиологические исследования занимают особое место. Такие исследования проводят с использованием показателей фоновой ритмики (активности) и вызванных потенциалов разных структур мозга, импульсных разрядов и мембранных потенциалов отдельных нервных клеток. Сопоставление полученных данных в сочетании с показателями условных рефлексов позволяет выявить некоторые сведения о механизмах, свойствах образования и осуществления временных связей.
Фоновая ритмика (активность). Непрерывно идущие колебания потенциалов мозга, обозначаемые как фоновая (спонтанная) или основная ритмика, меняют свой характер, отражая течение процессов высшей нервной деятельности. Это явление было обнаружено случайно. После того как несколько раз фотографировали с экрана осциллографа реакцию депрессии альфа-ритма (десинхронизации) электроэнцефалограммы (ЭЭГ) человека на вспышку света, достаточно было испытуемому услышать щелчок затвора фотоаппарата и в его электроэнцефалограмме появлялась эта реакция. Звук щелчка стал сигналом реакции десинхронизации (активации) ЭЭГ, которая приобрела условный характер, что стали использовать для изучения процессов высшей нервной деятельности. Другой реакцией основной ритмики электрической активности мозга, весьма успешно используемой для этой цели, было явление усвоения ритма сигнального раздражителя (рис. 13).
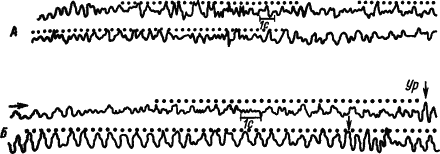
Рис. 13. Синхронизация потенциалов моторной коры мозга кролика ритмическими вспышками света, совпадающими по частоте с электрокожным раздражением лапы при выработке двигательного оборонительного рефлекса. А — до выработки условного рефлекса; Б — после 150 сочетаний (по М.Н. Ливанову, К.Л. Полякову):
в каждой паре записей нижняя продолжает верхнюю, точками обозначены вспышки света, Ур — условно-рефлекторное движение лапы
Дальнейшие исследования показали, что реакция десинхронизации электрокортикограммы (ЭКоГ) человека и животных, так же как реакция усвоения ритма, может отражать многие свойства нервных механизмов условно-рефлекторной деятельности. Так, во время генерализации условных рефлексов эти реакции возникают на обширной территории коры, охватывая области проекций и условного и безусловного раздражителей, а по мере специализации условных рефлексов сосредоточиваются на более ограниченной зоне, большей частью в двигательной области коры.
В процессе образования временной связи реакции синхронизации и усвоения ритмов раздражителя вначале формируются в корковых проекциях сигнала, затем в зоне проекции подкрепления, при дальнейшем упрочении условного рефлекса идут на убыль в зоне проекции сигнала, но долго сохраняются в зоне проекции подкрепления. При этом усвоение ритма происходит сначала в верхних, а потом в нижних слоях коры. Парная работа полушарий в формировании временных связей проявляется в том, что по мере выработки условного рефлекса ритмы сигнального стимула усваиваются вначале структурами верхних слоев коры полушария, контралатерального стороне подкрепления, затем они появляются в его нижних слоях и во всех слоях ипсилатерального полушария, потом они уменьшаются и исчезают всюду, кроме нижних слоев полушария, контралатерального стороне подкрепления (рис. 14).
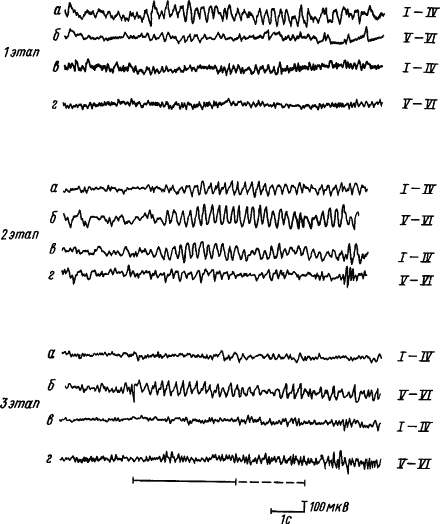
Рис. 14. Пространственная динамика усвоения ритма условного раздражителя в слоях симметричных пунктов коры больших полушарий головного мозга в процессе выработки условного рефлекса (по А.Б. Когану, Г.А. Кураеву):
I—VI — слои коры, а, б — запись ЭКоГ от полушария контралатерального относительно стороны подкрепления, в, г — то же, из ипсилатерального полушария; 1, 2, 3 — этапы выработки условного рефлекса (1 — первые сочетания, 2 — генерализованный условный рефлекс, 3 — упроченный специализированный), сплошной линией внизу обозначено действие условного стимула, пунктиром — безусловного
Разработка методов электроэнцефалографии с одновременной регистрацией ЭКоГ от десятков и сотен пунктов коры и использование электронно-вычислительных машин позволили выяснить роль синхронизации ритмов биоэлектрической активности в областях корковой проекции условного и безусловного раздражителей для образования временной связи между ними. Показано, например, что условные оборонительные движения лапы у кролика проявлялись тогда, когда коэффициент корреляции ритмов активности исследуемых корковых нейронов достигал достаточно высокого значения. Это подтверждено и для спонтанных движений. Функциональное значение пространственной синхронизации электрических потенциалов головного мозга было показано на многочисленных примерах. Показатели синхронизации также демонстрируют иррадиацию нервных процессов при начальной генерализации условного рефлекса, а по мере его закрепления и специализации нарастающую их концентрацию (рис. 15).