– Увеличение времени нахождения мостиков в ригорном – сцепленном не генерирующем силу состоянии (фаза V, VI);
– Увеличение времени нахождения мостиков в разомкнутом состоянии (фаза I, II).
Вспоминая аналогию с гребцами, скажем, что скорость движения лодки и развиваемая при этом сила тем выше, чем быстрее весла вытаскиваются из воды после гребка и чем меньше гребцы сидят с поднятыми веслами без дела.
Время нахождения мостика в ригорном состоянии зависит от того, как быстро АТФ вступит в контакт с головкой миозина. Как известно частицы вещества постоянно находятся в хаотическом движении (рис. 10). Предположим что r – некий радиус, ограничивающий область пространства вокруг головки миозина, при попадании в пределы которого молекулы АТФ становится возможным реакция, приводящая к отцеплению головки от актина.
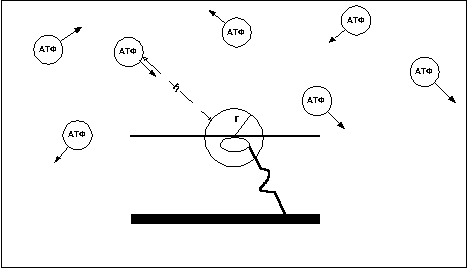
В этом случае время нахождения мостика в ригорном состоянии равно частному от деления расстояния (h) от мостика до ближайшей молекулы АТФ, движущейся в направлении мостика, на скорость движения молекулы (v). Среднее же время нахождения мостиков в ригорном состоянии равно средневзвешенному расстоянию между молекулами АТФ деленному на средневзвешенную скорость движения молекул.
Т=h/v
Естественно чем выше концентрация АТФ в мышце, тем меньше среднее расстояние между молекулами АТФ и головками миозина и тем меньше время нахождения мостиков в ригорном состоянии.
То есть сила волокна зависит от концентрации АТФ. При прочих равных условиях волокно с высокой концентрацией АТФ сильнее такого же волокна, но с меньшей концентрацией АТФ. Известно, что воспроизводство АТФ за счет креатинфосфата происходит почти мгновенно, поэтому можно сказать, что концентрация АТФ при работе мышцы зависит и от концентрации креатинфосфата. Таким образом:
Чем выше концентрация энергетических фосфатов (АТФ и креатинфосфата) в мышечном волокне, тем выше сила, развиваемая волокном.
После зависимости силы волокна от количества и поперечного сечения миофибрилл, влияние концентрации энергетических фосфатов на силу волокна является наиболее значимым. Именно по этой причине многие атлеты отмечают некоторую прибавку в силе, повышая уровень креатинфосфата в мышцах, принимая креатин, в качестве пищевой добавки.
Надо заметить, что зависимость силы волокна от концентрации энергетических фосфатов не является линейной. При высоких концентрациях, когда время ожидания контакта АТФ с головкой миозина станет меньше или сопоставимым со временем, необходимым для протекания химической реакции, приводящей к отделению головки миозина от актина, повышение концентрации АТФ и креатинфосфата будет мало влиять на скорость переключения мостиков и соответственно силу волокна. Зато низкие концентрации АТФ и креатинфосфата существенно снизят силу волокна, что может служить одной из причин отказа.
Рассмотрим второй фактор снижения силы сокращения – увеличение времени нахождения мостиков в разомкнутом состоянии (I, II фазы), что может происходить в результате снижения скорости гидролиза АТФ (снижения активности АТФазы – фермента ускоряющего реакцию гидролиза АТФ).
Расщепление АТФ и его воспроизводство за счет креатинфосфата вызывает накопление в мышце ортофосфорной кислоты, а гликолиз ведет к накоплению молочной кислоты. Активность АТФазы – фермента, благодаря которому происходит гидролиз АТФ, сильно зависит от кислотности среды. Как показывают эксперименты, максимум АТФазной активности достигается при среде близкой к нейтральной (рН=7), а при смещении среды мышцы в кислую сторону активность АТФазы падает, и при снижении рН среды до 5 АТФазная активность стремится к нулю (Богач П.Г. с соавторами, Поглазов Б.Ф). Таким образом, при накоплении кислых продуктов метаболизма АТФаза миозина постепенно снижает скорость гидролиза АТФ, и мостики теряют способность сцепляться с актином, при этом мышца снижает силу сокращения, несмотря на поступающий от мотонейрона сигнал.
Существует и еще как минимум две причины снижения силы мышц – при длительной работе накопление продуктов метаболизма тормозит процессы передачи сигнала от мотонейрона к волокну (Романовский Д.Ю.), а в центральной нервной системе развивается охранительное торможение.
То есть «отказ» мышцы является суперпозицией различных причин и доминирование одного либо другого фактора определяется интенсивностью работы мышцы.
Резкая активизация мышечной деятельности из состояния покоя требует такого же резкого увеличения скорости производства энергии.
Для достижения максимальной мощности основных источников воспроизводства энергии (гликолиза в быстрых волокнах и окисления в медленных) требуется время.
Скорость воспроизводства АТФ за счет гликолиза достигает своего максимума только через 20 —30 секунд после начала интенсивной работы.
Для достижения максимальной скорости окислительного процесса требуется гораздо больше времени, связано это в основном с необходимостью оптимизацией процессов доставки кислорода. Скорость окисления становится максимальной лишь через 1-2 минуты работы мышц, этот эффект наверняка известен вам под названием «второе дыхание».
Между тем мышца развивает максимальную мощность с первых же долей секунд после поступления команды к сокращению, гликолиз, в совокупности с окислением, не в состоянии обеспечить необходимую скорость воспроизводства АТФ для поддержания этой мощности. Приведение в соответствие скоростей расхода и воспроизводства АТФ во время работы мышцы идет по двум направлениям. Во-первых, постепенная активизация гликолиза и окисления увеличивает количество АТФ, синтезируемого в единицу времени за счет этих источников. Во-вторых, накопление продуктов метаболизма, в результате деятельности гликолиза и окисления, снижает активность АТФазы миозина и соответственно скорость расхода АТФ. Благодаря этим двум процессам скорости расхода и воспроизводства АТФ выравниваются, и в дальнейшем движение продолжается с постепенно снижающейся мощностью, но в состоянии равновесия между количеством синтезируемого АТФ и потребностями мышцы в энергии. Отказ же мышцы наступает не из-за окончания запасов АТФ, а из-за снижения сократительной способности мышц в результате накопления кислых продуктов метаболизма.