Зеркальные нейроны, как мы уже видели, не только коррелируют с сокращениями отдельных групп мышц, но и создают эквивалентность с конкретными двигательными актами и тем самым составляют то, что было названо "моторным словарем" (Rizzolatti and Gentilucci 1988). Очевидно, что множество данных об имитационном поведении (например, Meltzoff and Moore 1977) и такие простые факты, как способность младенцев улыбаться в возрасте 3 месяцев, задолго до того, как они смогут узнать себя в зеркале, предсказывают существование функционального уровня в Я-модели, который обеспечивает это отображение от визуального входа к имитационному моторному выходу, а также облегчает все те процедуры обучения, которые коренятся в способности к имитации, способности к выполнению намеченных, моторных самосимуляций. Однако, если применить к этой вновь открытой системе расширенный телеофункционалистский анализ, возникает интересная гипотеза, согласно которой она является рудиментарным элементом именно того механизма, который впоследствии позволил нам овладеть народной психологией и развить социальное познание. Это также имеет прямое отношение к философским теориям о том, что, собственно, значит обладать чем-то вроде народной психологии. Теория народной психологии анализирует нашу способность понимать ментальные состояния других людей как теоретическую деятельность, которая применяет теоретические умозаключения и концепции, будучи оторванной от воплощенного субъекта, то есть как чисто когнитивную деятельность. С другой стороны, симуляционная теория народной психологии - которая, очевидно, должна быть вариантом, которому отдает предпочтение предлагаемая здесь теоретическая модель - утверждает, что понимание ментальных состояний других людей основывается на бессознательной самосимуляции, внутренней, ненамеренной имитации, которая приводит к реальному "воплощению" ментальных состояний и поведенческих целей, являющихся целью этого вида репрезентативной деятельности. Я называю этот процесс "метаэмуляцией": Биосистема, обрабатывающая информацию, эмулирует саму себя, как в данный момент эмулирует другого агента. Предсказание, связанное с этой новой концепцией метаэмуляции, заключается в том, что физический субстрат и необходимый вычислительный инструмент для этого нового репрезентативного достижения идентичны определенным разделам человеческой Я-модели. Тот факт, что такое ментальное подражание другим когнитивным субъектам посредством процесса внутренней самосимуляции действительно существует, является, как отмечают Галлезе и Голдман, предсказанием теории симуляции, но не теории-теории (Gallese and Goldman 1998, p. 497). Та часть самомодели человека, которая реализуется зеркальной системой в основном бессознательно и автоматически, активирует в человеке-наблюдателе состояние, соответствующее состоянию наблюдаемого агента, что представляет собой внутренний тип имитационного поведения на функциональном уровне, метко названный (subpersonal) "попытка воспроизвести, подражать или выдать себя за психическую жизнь агента-мишени" (там же). Таким образом, управляемая извне активность этой части не-ПСМ может служить цели "ретродикции" психического состояния целевого лица, не только активируя моторную самосимуляцию у наблюдающего субъекта, но и одновременно активную репрезентацию цели. Обратите внимание, что функциональное прочтение ограничения свернутого холизма, обсуждаемого в разделах 3.2.4 и 6.2.4, делает такое предположение правдоподобным.
Что такое действие, в отличие от поведения, с точки зрения теории самомоделей? Действие - это поведение, вызванное внутренней моторной самосимуляцией, которая дополнительно сопровождается абстрактным представлением цели, представленным, во-первых, в неэгоцентрической системе отсчета, и, во-вторых, представлением фактического процесса выбора конкретного движения (например, Haggard and Eimer 1999), которое затем интегрируется в PSM, становясь, таким образом, "моим собственным намерением действовать". Функциональная связь, которую разделяют обе части сознательной моторной Я-модели, как канонические нейроны, так и зеркальные нейроны, заключается в том, что они легко связаны с аллоцентрическими репрезентациями целей. Цель другого агента распознается наблюдателем при отображении ее на общую моторную репрезентацию. Таким образом, то, что происходит, когда мы осознанно понимаем намерение другого воспринимаемого агента, может заключаться в обнаружении абстрактного свойства, а именно "моторной эквивалентности". Если это так, то степень сходства функциональной структуры Я-моделей двух классов систем определяет и степень успешности социального познания, успешности интерсубъективности, которая может иметь место между ними. Теория зеркальных нейронов для симулятивного чтения мыслей ценна тем, что открывает совершенно новую интерпретацию функции этой части Я-модели человека, а именно как "детектора намерений". Она помогает системе распознавать стимулы как волевые, определяя их цель в терминах внутренней самосимуляции (Gallese 2001). Еще один интересный аспект этого направления мысли заключается в том, что большинство функциональных аспектов человеческой Я-модели, обеспечивающих интерсубъективность и социальное сознание, явно неконцептуальны, дорациональны и дотеоретичны - это особый вид симулятивно-репрезентативного содержания, воплощенного в эгоцентрических аспектах моторного словаря. Те части Я-модели человека, которые можно описать как набор моторных схем, позволяют преобразовывать внешнее знание (представленное наблюдениями за моторным поведением других людей) во внутреннее знание (осознанное открытие того, что в себе самом такое поведение обычно обусловлено определенными целями) и тем самым генерировать глобально доступное знание о внутренних аспектах тех частей внешней реальности, которые на самом деле представлены другими агентами. Это знание - знание посредством воплощенной метаэмуляции.
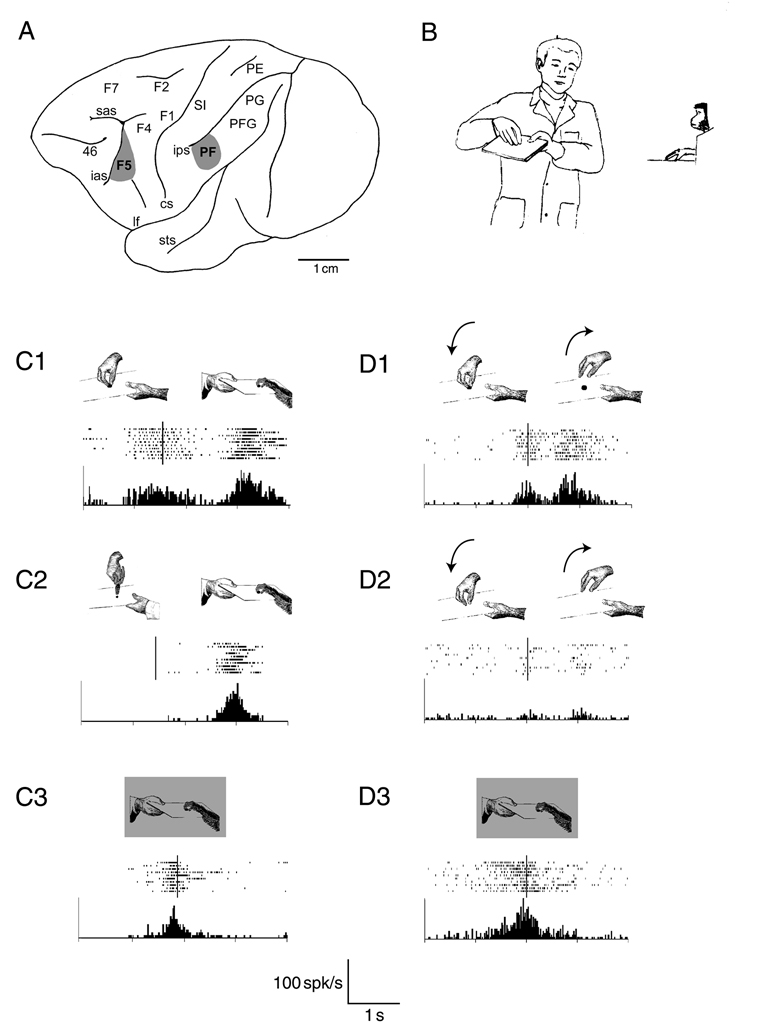
Чтение мыслей с помощью уже существующей внутренней самомодели: Обезьяна макака использует части собственной двигательной системы для мысленного моделирования человеческого агента, направленного в данный момент к конкретной цели действия. A, Боковой вид коры головного мозга макаки с изображением лобных и теменных областей. Лобные агранулярные области коры классифицированы в соответствии с Matelli et al. (1985). Задние теменные области классифицированы в соответствии с Von Bonin и Bailey (1947). Заштрихованные области обозначают сектора коры, в которых регистрировались зеркальные нейроны. Сокращения: cs - центральная борозда; ias - нижняя дугообразная борозда; ips - интрапариетальная борозда; lf - латеральная борозда; SI - первичная соматосенсорная область; sas - верхняя дугообразная борозда; sts - верхняя височная борозда. B, Иллюстрация экспериментальной ситуации для проверки визуальных свойств зеркальных нейронов. (Модифицировано из diPellegrino et al. 1992.) C, Пример зрительного и моторного ответов зеркального нейрона F5. Поведенческая ситуация, во время которой регистрировалась нейронная активность, схематично показана в верхней части каждой панели. В нижней части показаны растры и гистограммы относительных перистимульных ответов. C1 - обезьяне предъявлялся поднос с куском пищи; экспериментатор брал пищу в руки, а затем двигал поднос с пищей к обезьяне, которая брала его в руки. Сильная активация присутствовала во время наблюдения за хватательными движениями экспериментатора и во время выполнения того же действия обезьяной. Обратите внимание, что нейронный разряд отсутствовал, когда пища предъявлялась и двигалась к обезьяне. C2, Как и в C1, за исключением того, что экспериментатор захватывал пищу щипцами. Обратите внимание на отсутствие реакции, когда наблюдаемое действие выполнялось с помощью инструмента. C3, Обезьяна хватала еду в темноте. Растры и гистограммы выровнены (вертикальная полоса) по моменту, когда экспериментатор (C1 и C2) или обезьяна (C3) коснулись пищи. Абсциссы: время. Ордината: шипики/с. Ширина интервала: 20 мс. (Модифицировано из Rizzolatti et al. 1996). (D) Пример визуального и моторного ответа префронтального зеркального нейрона. D1, обезьяне был предъявлен поднос с куском пищи; экспериментатор схватил пищу и затем отпустил ее, отведя руку от пищи. Обратите внимание на сильный ответ во время наблюдения за действиями экспериментатора по захвату и отпусканию пищи. Нейрон не реагировал во время представления пищи на подносе. D2, Имитировалось то же действие, что и в D1. Обратите внимание, что в этом состоянии нейронный ответ практически отсутствовал. D3, Обезьяна схватила еду в темноте. Растры и гистограммы выровнены (вертикальная полоса) по моменту, когда обезьяна (D3) или экспериментатор касались пищи (D1) или подноса (D2). Все остальные условные обозначения как на рисунке C2. (Изменено из Gallese et al. 2002).
Важно отметить, что сознательное переживание того, что человек понимает текущие намерения и цели других людей, может возникнуть только в том случае, если внутренне представленная цель действий будет успешно приписана внешнему, наблюдаемому в данный момент агенту и если этот довольно сложный вид ментального содержания станет глобально доступным. Например, можно предположить, что особенно чувствительное или гиперактивное "устройство обнаружения агента" может привести к осознанному переживанию присутствия невидимого агента (Barrett 2000). Важный вопрос, конечно, заключается в том, могут ли самомодели, обладающие необходимыми функциональными свойствами, приводящими к низкоуровневому моторному резонансу и подражательному поведению в группах биологических организмов, существовать без сознательной способности к чтению мыслей.