Однако заманчивая перспектива связать положительные эмоции исключительно с парасимпатической системой, а отрицательные - с симпатической вряд ли соответствует действительности. Многочисленные исследования показывают, что между симпатическим и парасимпатическим отделами существуют гораздо более сложные отношения, чем прямая реципрокность. Нередко нарастание возбуждения симпатического отдела сопровождается одновременным нарастанием активности парасимпатических центров (Гельгорн, 1948, 1959, 1960; П. В. Симонов, 1964а, и др.). В структуре положительных эмоций мы обнаруживаем явно симпатические эффекты, а в структуре отрицательных - парасимпатические. Существует ли какая-нибудь закономерность в этих симпато-парасимпатических отношениях или при возникновении разных эмоций симпатические и парасимпатические реакции комбинируются в самой причудливой последовательности? По-видимому, определенная система все-таки существует.
Небольшой сдвиг вегетативного баланса в парасимпатическом направлении наблюдается при состояниях покоя, комфорта, расслабления. Симпатический сдвиг характерен для бодрости и оптимизма, однако дальнейшее нарастание симпатического тонуса, как правило, уже свидетельствует о напряженности, агрессии и тревоге. Значительное возбуждение парасимпатического отдела мы обнаруживаем при сильном страхе, депрессии и меланхолии (Гельгорп. 1901).
Обращает на себя внимание тот факт, что активно-оборонительные реакции (агрессия, ярость) расположены в "симпатической сфере влияния", а пассивно-оборонительные (страх, депрессия) - в "парасимпатической сфере". По данным И. Т. Джелиева, Н. И. Лагутиной и А. А. Фуфачевой (1963), активно-оборонительные реакции у обезьян сопровождаются тахикардией, а пассивнооборонительные - урежением сердечного ритма. В этом плане становится понятной и парадоксальная для "позитивно-негативной" концепции локализация центров страха в переднем отделе гипоталамуса наряду с реакцией агрессии при раздражении заднего отдела (С. А. Саркисов, 1964).
Рассматривая симпато-парасимпатические отношения при различных эмоциональных состояниях, мы обнаруживаем явное сходство с теми отношениями, которые складываются в вегетативной сфере по мере нарастания силы действующего раздражителя (П. В. Симопов, 1962). Вместе с тем в "ряду эмоций", о котором мы только что говорили, ость одно непонятное звено: преобладание парасимпатического отдела там наблюдается дважды в самом начале "ряда" (покой, комфорт, расслабление) и в самом конце (депрессия, сильный страх). Изучение закономерных изменений вегетативных реакций организма на возрастающий стимул убедило в том, что состояние первичного угнетения существенно отличается от вторичного, запредельного по своей природе. Первичное торможение возникает задолго до того, как нервные структуры исчерпают свои функциональные возможности. При действии достаточно сильного стимула первичное торможение (мы назвали его превентивным) исчезает и сменяется возбуждением. Условия возникновения и биологическая роль превентивного торможения представляют частный случай принципа гиперкомпенсации, о котором мы подробно говорили в предыдущей главе. Весьма вероятно, что преобладание парасимпатической системы при комфорте и удовлетворении реализуется на фоне первичного торможения, а парасимпатические эффекты депрессии и страха выступают па фоне запредельного. Пока трудно сказать, чем отличаются нейрофизиологические механизмы превентивного торможения от механизмов запредельного, с какими структурами связаны эти два вида центрального угнетения. По-видимому, в осуществлении первого из них решающая роль принадлежит кортикальным тормозным аппаратам и неспецифическим ядрам таламуса (Жуве, 1950; А. И. Ройтбак, 1959, 1962). Запредельное торможение связано с более глубокими физико-химическими сдвигами в нервных клетках, в частности с повышением сорбционных свойств клеточной протоплазмы (С. Н. Романов, 1956; Д. Н. Насонов, 1959). Возможно, что при запредельном торможении корковых клеток усиливается тормозящее влияние ретикулярной формации нижнего отдела мозгового ствола. По мнению Моруцци (1902), ото тормозящее влияние сдерживается корой больших полушарий. Мы ужо говорили, что возбуждение симпатического отдела вегетативной нервной системы в большинстве случаев сопровождается активирующим действием ретикулярной формации.
На рис. 3 схематически изображено участие симпатического и парасимпатического отделов вегетативной нервной системы, а также механизмов возбуждения, превентивного и запредельного торможения в осуществлении различных эмоциональных состояний. Схема показывает, что отношения между симпатической и парасимпатической активностью по сводятся к прямой реципрокности. Возбуждение симпатического отдела всегда сопровождается возбуждением парасимпатического отдела, однако "удельный вес" каждой функциональной составляющей специфичен для той или иной группы эмоций. Нельзя не обратить внимания на сходство между схемами рис. 3 и фазными изменениями вегетативных реакций организма по мере нарастания возмущающего воздействия (П. В. Симонов, 1964). Впрочем так ли уж неожиданно это сходство? Рассматривая группы эмоций на рис. 3, мы видим, что для эмоций, расположенных в правой частп схемы, характерны большая степень потребности и больший дефицит информации, чем для состояний, перечисленных слева (согласно формуле, величина эмоции прямо пропорциональна и потребности, и дефициту). Специального анализа требует только место положительных эмоций (второй столбик слева). Их размещение на схеме хороню согласуется с результатами экспериментов.
Мы не раз отмечали, что положительные эмоции по своим вегетативным проявлениям уступают сдвигам в организме, связанным с отрицательными эмоциями, хотя и требуют большей степени напряжения, чем состояние покоя. Но ведь положительные эмоции возникают при избытке информации и но этому признаку должны располагаться слева от покоя. Возникшее противоречие устраняется "формулой эмоций". Наши схемы отражают величину эмоционального напряжения, а не ее знак. Величина Э может достигать значительных размеров и при Н больше С, и при С больше Н. Впрочем при положительных эмоциях возбуждение практически не бывает чрезмерным: науке но известны нервные расстройства от слишком большой радости. Вся клиника неврозов базируется на отрицательных эмоциях, и никто еще не описал психосоматических (кортиковисцеральных) заболевании на почве счастья, удовлетворения и оптимизма. Только в психиатрии мы встречаем патологическое возбуждение позитивных структур (мания величия, эйфория), которое, по-видимому, возникает вторично в результате нарушения высших контролирующих аппаратов.
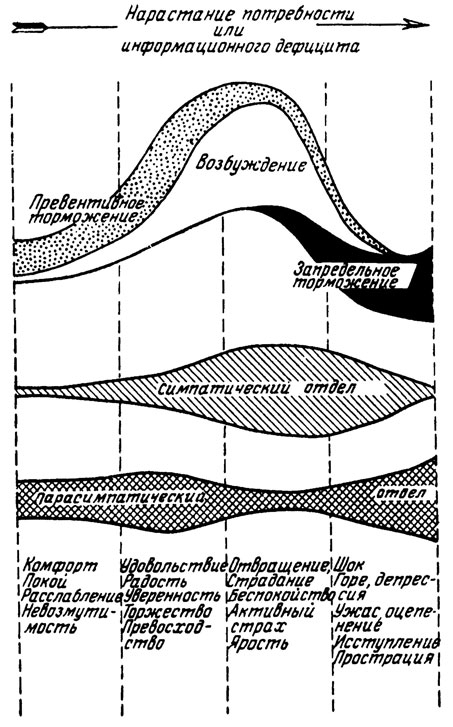
Рис. 3. Схема участия механизмов превентивного торможения, возбуждения и запредельного торможения, а также симпатического и парасимпатического отделов вегетативной нервной системы в реализации различных эмоциональных состояний
Итак, мы рассмотрели пути и закономерности реализации эмоциональных состояний. Теперь необходимо перейти к вопросу о механизмах, которые запускают ту или иную эмоциональную реакцию. Что касается потребностей, то в соответствующей главе были приведены данные о центрах голода, насыщения, жажды, полового влечения и т. д. Но где определяется дефицит информации? Откуда поступают команды к исполнительным центрам разнообразных эмоций?
По мнению И. С. Беритова, регуляция эмоций сосредоточена в древней коре. Именно палоокортекс производит интеграцию активности сомато-вегетативных центров гипоталамуса, центров внешнего выражения эмоций в полноценную эмоциональную реакцию высших позвоночных животных. Среди образований древней коры исследованиями последних лет особое место отводится гиппокампу. Было отмочено, что при решении животным какой-либо задачи (например, поиск пищи в лабиринте) электрическая актин ность гиппокампа усиливается. Она нарастает при ошибках, по исчезает в момент достижения цели (Эйди, 1900-1961; Холмс и Эйди, 1960). Электрическое раздражение гиппокампа ведет к нарастанию ошибок, которые перестают коррегироваться (Прибрам, 1960). Эти экспериментальные факты дали основание рассматривать гиппокамп в качестве аппарата сличения ожидаемого эффекта с реально достигнутым, который П. К. Анохин называет "акцептором действия" (А. Р. Лурия, 1963). Вместе с тем трудно допустить, чтобы сличение происходило в самом гиппокампе. Все данные современной физиологии свидетельствуют о том, что органом накопления индивидуального опыта, нервным устройством замыкания и хранения условных связей является новая кора больших полушарий. Как же тогда понять своеобразие электрической активности гиппокампа? Прежде всего напомним, что гиппокамп относится к числу образований, связанных с отрицательными эмоциями ярости и страха (И. С. Беритов, 1961; Икеда Терутика, 1961; Дельгадо, 1962; Монье-Дюман, 1963 и др.). Неудачи животного при решении задачи и возникающий при этом дефицит информации ведут к развитию отрицательного состояния, которое реализуется с участием гиппокампа. Таким образом, вовлечение гиппокампа в реакцию можно представить себе как явление вторичного порядка, отнюдь не обусловленное процессами сличения в самих гиппокампальных структурах.