Зерно полученное от прививки, изучалось А. А. Шмуком. Исследования Шмука показали, что трифруктозан, являющийся специфическим углеводом зерна ржи в отличие от зерна пшеницы, в контрольном материале отсутствует. Пять граммов муки привитой пшеницы дали 0,14 г трифруктозана.
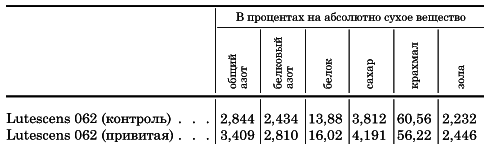
Согласно сообщению Писарева, клейковина, выделенная из муки контрольной пшеницы, имела обычную светлорозовую окраску; клейковина же подопытного варианта имела цвет ржаного теста (темнобурая окраска).
Писарев проводил серию опытов по скрещиванию привитых пшениц Lutescens 062, гибрида 170 и Авроры с рожью. Во всех случаях он получил закономерный эффект. Так, например, скрещивание обычных (контрольных) растений Lutescens 062 с рожью дало всего 4,3 % завязавшихся зёрен, привитые же растения этих компонентов дали 25 % зерна. Соответственные показатели по сорту гибрида 170 равнялись 3,4 и 11,5 %, по сорту Аврора — 2,8 и 19,2 %.
Писарев приходит к следующему выводу:
«Таким образом, сдвиг биохимической характеристики в яровой пшенице под влиянием пересадки её зародышей на эндосперм яровой ржи в сторону последней повлиял в значительной степени и на повышение процента гибридного зерна в межродовом скрещивании».
Писарев провёл ещё один любопытный опыт, основанный на вегетативном сближении двух нескрещивающихся родов — Triticum vulgare и Elymus arenarius.
В основу была положена та же методика. По заявлению Писарева «лучшие результаты получались в тех случаях, когда и материнское и отцовское растения были «привиты» на эндосперм компонента. Так, яровая пшеница 1803, трижды выросшая на эндосперме Е. arenarius 19, при опылении пыльцой последнего дала 1,55 % завязавшихся зёрен. В том случае, когда пыльца была взята с элимуса, выросшего на эндосперме пшеницы 1803, процент удачи равнялся 7,5; яровая пшеница Prelude, несмотря на двухкратную прививку на эндосперм элимуса, при скрещивании дала 0,0 %, но при опылении элимусом, выросшим на эндосперме пшеницы, процент зёрен был уже 3,1. Гибридная пшеница ВЕП2, дважды привитая на элимусе, при опылении пыльцой обычного элимуса, дала 0,4 % зёрен, а при использовании пыльцы «привитого» элимуса — 1,7 % и т. д.».
В экспериментах Писарева мы видим ясные и чёткие доказательства силы мичуринского метода и при вегетативной гибридизации злаковых. Как доказали И. В. Мичурин и Т. Д. Лысенко, основой этого метода является изменение типа обмена веществ в сторону соответствующего компонента. Но Писарев не может согласиться с такого рода толкованием. По своим теоретическим взглядам он не может, вернее, не хочет понять смысла ведущего звена в цепи мичуринских работ, и основанием для его опытов поэтому и служат не мичуринское учение о развитии живого, а «последние исследования Шандера о роли фитогормонов, заложенных в эндосперме зерна злаковых и их влиянии на развитие как зародыша, так и взрослого растения» (Писарев, 1944).
Итак, сама наследственная субстанция — ген неизменён, но продукт гена — геногормон, фитогормон может изменить ген привоя, притом соответственно, адэкватно.
Трудно усмотреть логичность в такого рода «логике».
Каким же образом представители «хромосомной теории наследственности» объясняют неопровержимые факты глубоких взаимовлияний между двумя привитыми компонентами — факты, полученные их собственными руками?
И здесь формальная логика приходит на выручку. Морганисты считают, что такого рода явление происходит, в основном, под влиянием веществ, выделяющихся из ядра клетки, пытаясь этим самым экспериментально обосновать теорию монополии (преимущественной роли) ядра в наследственности.
Особо удачным доказательством этого положения морганисты считают эксперименты Геммерлинга (1935) с одноклеточной водорослью ацетабулярией (Acetabularia), которыми, по их представлениям, «с определённой ясностью показана роль ядра в определении видовых особенностей организма» (М. Л. Бельговский, В. С. Кирпичников, А. А. Прокофьева-Бельговская, 1940).
Существо опытов Геммерлинга заключается в том, что безъядерные участки A. mediterranea он приращивал к содержащим ядро кускам другого вида A. Wettsteini. В некоторых случаях участки mediterranea полностью развивались по типу Wettsteini, в других — сращённые организмы имели признаки обеих форм. Вот как описывает эти опыты Геммерлинг: «Из безъядерных трансплантантов от mediterranea, посаженных на содержащий ядро ризоид Wettsteini, 23 приняли свойства Wettsteini. Здесь принимаются во внимание только те трансплантанты (16), из которых образовалась шляпа Wettsteini. В двух случаях в шляпе возникли также цисты Wettsteini, из которых вылупились гаметы и цисты того же вида. Особенно один результат показывает, что весь трансплантант развился в систему Wettsteini: пересаженный стебель от mediterranea сильно сократился, что является свойством, характерным для более мелкой Wettsteini. В ряде случаев… шляпа типа mediterranea возникла, как первое образование, вскоре после сращивания. Но в 6 случаях сначала возникли образования типа mediterranea (типичные мутовки и возможно недоразвитые шляпы), но затем шляпа Wettsteini. Возникновение образований mediterranea основывается на действии ядра Wettsteini. В этих случаях пересаживалась передняя часть mediterranea, содержавшая уже больше или меньше специфических виду формообразующих веществ. Они пошли в ход в первую очередь и, как и следовало ожидать, индуцировали образования mediterranea; лишь затем подействовало ядро Wettsteini».
На наш взгляд опыты Геммерлинга с одноклеточными водорослями являются замечательным доказательством, во-первых, адэкватности влияния подвоя на привой и, во-вторых, устойчивости наследственности безъядерного куска привоя, благодаря которой частично проявляются и сохраняются видовые особенности.
Безусловно, весь этот вкратце изложенный экспериментальный материал противоречит учению менделистов о независимом развитии прививочных компонентов, показывает несостоятельность хотя бы утверждения, что взаимовлияния между привитыми растениями «никогда не выходят за пределы нормы реагирования данного генотипа», что они представляют просто «количественные изменения уже имеющегося признака» (Лусс, 1935).
Не выдерживают никакой критики категорические утверждения менделистов о том, что при трансплантациях не изменяется окраска. Не лишнее будет подтвердить документально это утверждение из упомянутой работы Лусса.
«Практика с несомненностью и на громадном количестве примеров показывает, — пишет Лусс, — что при прививке разноокрашенных растений друг на друга они своей окраски обычно не изменяют; пигментированные ткани никогда не передают своей окраски соседним зелёным тканям своего компонента, как бы они близко друг к другу ни были расположены».
Приходится поражаться тому, как часто представители менделевско-моргановского направления в генетической науке ошибаются в своих выводах и утверждениях. При этом нельзя не подчеркнуть, что эти ошибки, как правило, касаются самых важных вопросов, по которым эти учёные ведут борьбу с мичуринской генетикой, и вскрываются представителями их же направления.
Не успел Лусс выступить с цитированным выше утверждением об отсутствии глубокого влияния подвоя на привой при прививках, как ряд других исследователей-морганистов на таком генетическом объекте, как дрозофила, и других организмах показали глубокое взаимодействие между компонентами и передачу специфических веществ от одних тканей другим.
Как известно, дрозофила относится к насекомым с полным внутренним превращением. Цикл развития дрозофилы от яйца до взрослой мухи протекает девять суток. Развитие от начала дробления оплодотворённого яйца до вылупления личинки занимает 20–22 часа и по 4 суток приходится на стадию личиночную и куколки. Окукливание личинки означает начало полного превращения. За исключением гонад и нервной системы, все личиночные органы разрушаются, заменяясь дефинитивными органами.