Для сусликов и сурков характерно латентное течение инфекции во время зимней спячки. У тарбаганов это было доказано еще в 20-х годах Wu Lien-Teh и R. Pollitzer [цит. по Wu Lien-Teh et al., 1936]. Исследователи установили, в частности, что во время спячки сурков чумной микроб сохраняется без потери вирулентности на месте введения и/или в регионарных лимфатических узлах. Через некоторое время после пробуждения у животных может развиваться острый процесс, сопровождающийся бактериемией. Однако нечувствительность спящих сурков к чуме не является абсолютной, поскольку они все-таки могут погибать от инфекции, а иногда на коже спящих сурков в местах заражения развиваются язвы, инфицированные чумным микробом, что не свойственно бодрствующим животным [Ралль Ю. М., 1965].
Латентная инфекция у сурков становится явной весной, после пробуждения. Аналогичная картина наблюдается у сусликов: латентное течение во время спячки с генерализацией процесса и гибелью отдельных зверьков после пробуждения [Гайский Н. А., 1925].
Описание чумы у сурков и сусликов будет неполным, если не упомянуть еще одну особенность патогенеза, а именно вариабельность процессов в активные периоды жизни грызунов. Так, описывая одну из эпизоотий, И. С. Тинкер [1940] указывал, что в ходе первого месяца (июль) эпизоотии смертность сусликов была очень высокой и обнаружение их трупов в степи в состоянии сильного окоченения были обычным явлением. Однако ни бубонов, ни видимых изменений во внутренних органах при этом у них не находили, хотя в отпечатках из органов обнаруживалось огромное число микробов. К середине лета у сусликов стали чаще обнаруживаться крупные бубоны (размерами до лесного ореха), увеличенная селезенка и некротические изменения в печени, но выделить возбудителя у таких сусликов было трудно, а в некоторых случаях и невозможно.
Существенные различия в патологоанатомической картине от весны к осени отмечаются также у больных сурков. Весной чаще встречаются остросептические формы заболевания с большим числом бактерий во всех органах и крови, а к концу лета нарастают продуктивные изменения и резко падает вероятность выделения чумного микроба [Петрунина О. М, 1951а].
Иная ситуация наблюдается у песчанок.
Полуденные песчанки активны круглый год. В соответствии с этим зараженных зверьков и их блох обнаруживают постоянно, но все же чаще в весенне-летний и осенне-зимний сезоны. Подобную закономерность, т. е. двугорбый характер эпизоотий, В. Н. Федоров и соавт. [1955] связывали с сезонными изменениями активности полуденных песчанок, возможными изменениями их физиологического состояния, а также с влиянием сезонных условий на видовой состав и активность блох, паразитирующих на грызунах. В известной мере это мнение основывалось на экспериментальных данных, опубликованных В. Н. Лобановым и В. Н. Федоровым еще в 1938 г. Согласно им, в период с июля по октябрь большинство полуденных песчанок погибало от острых генерализованных форм чумы, а с апреля по июль у них отмечались локализованные формы с явной тенденцией к разрешению процесса, причём выделить культуру Y. pestis от таких животных не удавалось даже через биопробы.
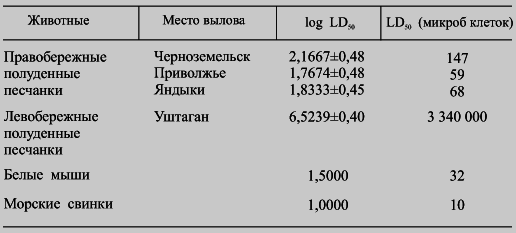
Необходимо напомнить, что полуденные песчанки являются носителями чумы в песчаном волго-уральском очаге, тогда как на правом берегу Волги, в степном очаге северо-западного Прикаспия, в поддержании чумы они никакой роли не играют [Ралль Ю. М., 1965]. Объяснение столь парадоксальной ситуации было найденоЕ. С. Бирюковой [1960], которая впервые в 1955–1957 гг. подметила различия в чувствительности к чуме полуденных песчанок, обитающих на левом и правом берегах реки Волги: чувствительность к чуме первых оказалась на несколько порядков выше, чем вторых. При этом очень важно отметить, что эти различия чувствительности двух популяций полуденных песчанок выявлялись не только при искусственном заражении, но и в случае укусов их инфицированными блохами [Ю. М. Елкин, 1960].
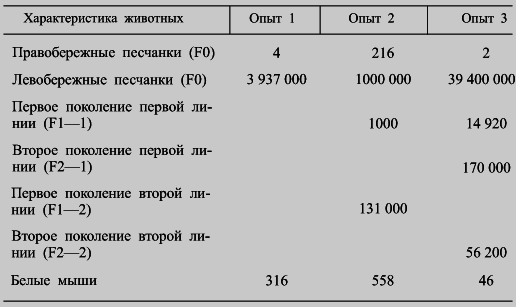
Вопросами чумы у полуденных песчанок много занимались М. И Леви и его сотрудники, подтвердившие первоначальное наблюдение Е. С. Бирюковой (табл. 6). При этом были установлены существенные различия в патогенезе чумы у право- и левобережных песчанок, подтверждающие гипотезу о том, что основным носителем чумной инфекции в природе являются «не столько высоко восприимчивые к чуме, сколько малочувствительные грызуны с высоким уровнем бактериемии». Кроме того, М. И. Леви, одному из немногих, удалось показать, что «резистентность к чумному микробу передается по наследству» (табл. 7). Небезынтересно, что отмеченные выше различия в чувствительности полуденных песчанок к чуме не распространяются на их чувствительность к псевдотуберкулёзу, туляремии и бруцеллезу, т. е. достаточно специфичны.
Не меньший интерес представляют сведения о заболеваемости чумой больших песчанок, обитающих на огромных просторах равнинного среднеазиатского очага. Здесь сохраняется в общем тот же характер эпизоотий, как и в волго-уральском очаге. Зараженных чумой больших песчанок можно обнаружить в любой сезон, но пики эпизоотий также приходятся на весну и осень. Зимние спады объясняют резким уменьшением активности песчанок, тогда как подъемы связывают с повышением подвижности грызунов и увеличением численности блох рода Xenopsylla. Однако число зараженных особей в популяциях песчанок в различные сезоны бывает неодинаковым, что опять-таки объясняется неодинаковой подвижностью грызунов в различное время года. Обращают на себя внимание отличия в инфекционной чувствительности отдельных возрастных групп, разброс индивидуальной чувствительности и зависимость её от сезона. Зарегистрированы и различия чувствительности к чуме у больших песчанок в разных популяциях. Например, зверьки из северо-восточного Прикаспия по сравнению с песчанками из Туркмении и особенно из Или-Каратальского междуречья оказались более чувствительными к инфекции [Наумов Н. П. и др., 1972]. Интересно, что в период затухания эпизоотий увеличивается процент затяжных и хронических форм чумы, столь характерных для больших песчанок. При этом часто выделяются авирулентные штаммы чумного микроба. Относительно вялое течение чумы с тенденцией к переходу в хронические формы подтверждено в экспериментах О. М. Петруниной [1951б] и А. А. Левиной [1960]. В опытах последней некоторые песчанки жили по несколько месяцев после заражения, но септицемия у них отмечалась очень редко, да и то в тех случаях, когда они погибали не позднее 35 сут.
Относительная устойчивость к чуме наряду с индивидуальными колебаниями чувствительности и тенденцией к затяжному течению присуща также краснохвостой и некоторым другим видам песчанок [Ралль Ю. М., 1965]. В отличие от них такие песчанки, как тамарисковая и Виноградова или Tatera brantsi, обладают высокой чувствительностью, на которую мало влияют климатические и другие факторы. В частности, тамарисковая песчанка легко вовлекается в эпизоотии среди грызунов иных видов и быстро погибает от острых септических форм чумы, что не раз позволяло использовать их в качестве хороших объектов для биологических проб на чуму.
В отличие от зимоспящих грызунов и песчанок представители семейства заячьих (пищухи стали объектами внимания относительно недавно.
В ходе эпизоотий среди сурков и сусликов в Забайкалье часто находили трупы пищух, павших от чумы; при вскрытии у них обнаруживали картину сепсиса. Это побудило Н. А. Гайского и Н. Д. Алтареву [1944] провести соответствующие исследования. Используя разные заражающие дозы, они установили, что от чумы погибает около 50 % даурских пищух. Тем не менее их все еще продолжали относить к числу второстепенных носителей [Некипелов Н. В., 1959б]. Однако отношение к пищухам изменилось, когда в 1955 г. в относительно спокойном районе Северо-Западной Монголии, недалеко от границы с Горным Алтаем, среди пищух была зарегистрирована эпизоотия чумы, а в 1961 г. она появилась и здесь. Сначала эпизоотию среди монгольских пищух в Горном Алтае рассматривали как результат заноса чумы из Монголии [Домарадский И. В. и др., 1963а], но в последующем очаг чумы на северном и южном склонах хребта Сайлюгем стали относить к издавна существующим очагам северо-восточной окраины центрально-азиатской «чумной зоны» [Голубинский Е. П. и др., 1987].