Приведенные данные не должны однако расцениваться как попытка свести на нет роль гуморальных факторов в защите от чумы. Не подлежит сомнению их опсонизирующая роль, на что указывалось выше, а также превентивное, хотя и слабо выраженное действие специфических антител [Meyer K. 1950]. Так или иначе, но антитела безусловно являются показателем иммунитета против чумы, что доказано многолетней практикой применения серологических методов для диагностики чумы в полевых условиях и даже у людей [Butler T., 1983]. При этом особое значение имеет выявление IgM (показателя недавнего контакта с источником чумы [Наумов А. В., Самойлова Л. В., 1992].
К сожалению, за последние годы иммуногенезу как таковому стали уделять меньше внимания, чем изучению проблемам патогенеза и роли в нем продуктов «основных» плазмид, хотя весьма вероятно, что в последующем это окупится сторицей. Ведь соответствующие вопросы тесно переплетаются; они как две стороны одной и той же медали. Кстати, в процессе изучения патогенеза открываются новые аспекты антигенной структуры чумного микроба и становятся более понятной роль уже известных.
Одной из характерных черт иммунитета к чуме является то, что его острие направлено против возбудителя, хотя в клинической картине чумы доминируют симптомы интоксикации. Помимо сомнительной эффективности иммунизации животных различными токсическими препаратами из Y. pestis [Pollitzer R., 1954], в пользу этого говорит низкое содержание или полное отсутствие антитоксина в сыворотке крови лиц, переболевших чумой [McCrumb F. R. et al., 1955; Warren J. et al., 1955; Payne F.E. et al., 1956]. К этому добавим, что при чрезмерных дозировках антибиотиков некоторые больные погибают от интоксикации, вызванной интенсивным распадом бактерий [Bahmanyar M., Cavanaugh D. C., 1976], несмотря на то, что антитела к возбудителю появляются уже с 7-го дня заболевания [McCrumb F. R. et al., 1955]. Возможно, борьба организма с токсемией при чуме осуществляется непрямым путем за счёт предотвращения размножения бактерий до критического предела. Однако нельзя исключить и другой возможности, а именно, что «истинный» токсин образуется только in vivo, и, не имея его мы не можем выявить антитоксин. Но тогда бы сыворотка крови переболевших должна была бы обладать лечебными свойствами.
Сейчас трудно сказать, какому из многочисленных антигенов чумного микроба [Lawton W. D. et al., 1960]. принадлежит решающая роль в иммуногенезе. Дело осложняется еще тем, что реакция различных видов животных на один и тот же антиген неодинакова [Burrows T., 1963]. В итоге мы не всегда можем быть уверены в том, что данные полученные на животных, безоговорочно приложимы к объяснению процессов, происходящих в организме человека.
Как указывалось, развитие иммунитета против чумы, как и против многих других инфекций, сопровождается сенсибилизацией организма, которая может быть выявлена с помощью специфических аллергенов [Коробкова Е. И., 1955]. Положительные реакции на аллергены появляются в первые недели после иммунизации, причём у морских свинок в те же сроки, что и иммунитет, или позже на 1–2 дня [Павлова Л. П., 1964; Павлова Л. П. и др., 1964]. У людей сроки появления гиперчувствительности замедленного типа (ГЗТ) зависят от способа и кратности введения живой вакцины. На основании этих и последующих данных было сделано заключение о целесообразности применения аллергенов для оценки эффективности вакцинации и даже для «выявления сенсибилизации к чуме в природных очагах у малого суслика» [Тараненко Т. М., 1988]. Укажем также, что Л. П. Павловой была установлена обратная связь между интенсивностью реакции на аллерген (пестин (и исходом заболевания морских свинок при заражении их чумой; отрицательная или слабая реакция служила плохим прогностическим признаком, Если бы это удалось подтвердить на людях, то отпали бы сомнения в наличии корреляции между ГЗТ и иммунитетом к чуме, которые высказывали Т. Burrows [1963], B. J. Hurtrel и соавт. [1981].
Вскоре после открытия чумного микроба были предложены различные убитые вакцины и отдельные компоненты микробной клетки [Домарадский И. В., 1966; Николаев Н. И., 1968; Wu Lien-Teh et al., 1936; Pollitzer R., 1954]. Однако при тщательном изучении всех этих препаратов ни одному из исследователей не удалось получить напряженного иммунитета у морских свинок. За редким исключением они были недостаточно эффективны и в полевых условиях, в очагах чумы. К тому же убитым вакцинам подчас была присуща высокая реактогенность. Поэтому давно уже стали проводиться поиски путей создания живых вакцин.
Уже W. Kolle и R. Otto [1903] показали, что только живые вакцины надежно предохраняют лабораторных животных от чумы. Н. Н. Жуков-Вережников [1940] при заражении морских свинок большими дозами вирулентной культуры (100 DLM) установил, что если убитые вакцины лишь удлиняли сроки жизни животных, то живые способствовали их выживанию. В работах других авторов [Покровская М. П., 1934; Коробкова Е. И., Крайнова А. Н., 1939; Коробкова Е. И., 1956; Е. Jawetz, К. Meyer, 1943, 1944] также была показана высокая эффективность живых и слабая убитых вакцин, однако сравнение их в очагах чумы оказалась далеко не легким делом. Даже в тех случаях, когда одних членов семьи прививали, а других оставляли непривитыми, трудно было установить, кто из них подвергался большей опасности заражения. Вследствие этого для оценки эффективности вакцинации избрали такой показатель, как снижение смертности населения [Николаев Н. И., 1964]., хотя и он не был бесспорным [Girard G., 1963].
К настоящему времени известно большое число вакцинных штаммов, полученных: путем селекции культур, в течение длительного времени пересевавшихся на питательных средах(таким образом были получены штаммы Tjiwidei (TJW), EV, 1 и17); культивированием при неблагоприятных условиях(шатамм М-11-40); воздействием бактериофагов (штаммы АМП, ЖВР, 46S).
Сведения о других методах получения вакцинных штаммов имеются в работах Коробковой Е. И. [1964], Николаева Н. И. [1964] и R. Pollitzer [1954],
Из всех штаммов к сегодняшнему дню лучше всего зарекомендовали себя TJW и EV.
Применяя штамм TJW на Яве, L. A. Otten [1941] установил, что среди 37436 привитых людей смертность от чумы была 1,01:1000 человек, а среди непривытых (4,75:1000.
Гораздо лучшие результаты были получены при использовании штамма EV, которое началось на Мадагаскаре. Там G. Girard [1963] в 1933 г. привил этим штаммом 13 тыс. человек. Вакцинация прошла без всяких осложнений и эффективность её, хотя и не была абсолютной, но во много раз превзошла таковую убитых вакцин, применявшихся на Мадагаскаре в течение предыдущих 10 лет. Продолжая прививочную кампанию, G. Girard руководствовался принципом массовости и, следовательно, необходимости сравнения с контрольными группами отпала. С ноября 1935 г. по ноябрь 1938 г. на Мадагаскаре было привито и ревакцинировано свыше 2 млн. человек. В результате заболеваемость чумой при неизменившейся эпизоотологической обстановке с 3045 случаев в 1935 г. снизилась до 596 в 1938 г. (рис. 14).
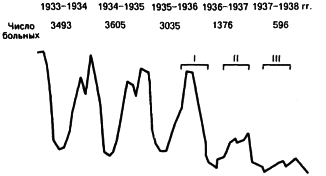
Рис. 14. Кривая заболеваемости чумой на Мадагаскаре до использования штамма ЕV и по ходу прививочных кампаний (I, II, III) [Коробкова Е. И., 1956].
Не менее убедительные данные об эффективности штамма EV были получены в Бельгийском Конго [Devignat R., 1949] и в Южной Африке [Grasset E., 1946].
По наблюдениям Н. И. Николаева [1968], в Ванемяо (Внутренняя Монголия) в 1945 году заболеваемость чумой среди 19780 привитых вакциной EV была 2,5 на 10 000, а среди непривитых 288:10 000.
В. П. Смирнов [1962] изучал эффективность сухой живой вакцины EV в Монголии, вводя её подкожно или конъюнктивально. По его данным, из 7885 привитых подкожно лёгочной чумой заболело 6 человек, а из 11 6333 человек, привитых через конъюнктиву, 2 человека (1 — бубонной и 1 — лёгочной), среди 31 878 непривитых было 69 случаев, из них 48 лёгочной чумы.