Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пелагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодпой Атлантике, например, в последние годы резко увеличилась численность серого спинорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus (рис. 15). Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Их выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
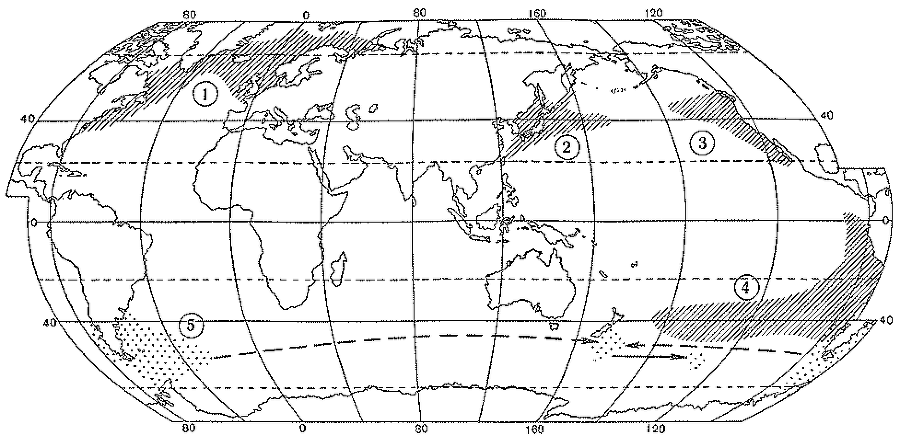
Максимально расширенные ареалы псевдонеритических рыб - атлантической сельди (1), сардины иваси и дальневосточной скумбрии (2), калифорнийской (3) и перуано-чилийской ставриды (4) и возможные пути расселения южной путассу (5)
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
В двух последних случаях вид может рассматриваться как временно или постоянно обосновавшийся в том или ином конкретном участке открытого океана. Примеры, иллюстрирующие сказанное, ограничиваются, к сожалению, данными за самые последние десятилетия, так как имеющиеся ряды наблюдений очень коротки. Большая часть этих примеров относится к Северной Пацифике.
Известно, что в период высокой численности японского анчоуса и японской скумбрии в 50-е годы оба вида были весьма обычными и в открытых водах северо-западной части Тихого океана, в которых предполагалось даже существование их особых «стад» — в действительности, по всей вероятности, псевдопопуляций. В этом же районе в период высокой численности сардины иваси (Sardinops sagax melanosticta) в 30-е годы ее ареал расширялся вплоть до южной части Берингова моря (наблюдения в открытом океане отсутствовали), а современный подъем численности, начавшийся в 70-е годы (за первые 5 лет роста запас возрос не менее чем вдвое), привел к расширению области распространения в восточном направлении вплоть до центральной части океана, где, судя по некоторым сведениям, также образовывались временные псевдопопуляции. У японской скумбрии, как показано В. А. Беляевым, усиление воспроизводства на нерестилищах в прибрежной зоне Японии в середине 70-х годов привело к заселению океанических участков и формированию в районе Северо-Западного подводного хребта (гора Кинмей и прилегающие воды) зависимой популяции псевдонеритического типа: в 1978 — 1981 гг. скумбрия наблюдалась там на всех фазах жизненного цикла (икра, личинки, мальки, сеголетки).
Интересный материал для размышлений представляют сведения по биологии и распространению двух восточнотихоокеанских видов ставрид из группы Trachurus picturatus. Размножение калифорнийской ставриды (Т. symmetricus simmetricus) было отмечено по нахождениям икры и личинок в зоне 40-х параллелей северного полушария в 1955г. в удалении до 1100 миль, а в 1972г. — до 600 миль от берегов Северной Америки. Аналогичным образом нерест перуанско-чилийской ставриды (Т. s. murphyi) в 40-х широтах южного полушария отмечен в 1985г. С. А. Евсеенко на огромном расстоянии (около 3000 миль) от побережья Чили. Никаких свидетельств в пользу результативности такого нереста, впрочем, не существует, так как в этих океанических участках — у крайних границ своего распространения на запад — оба вида представлены, судя по имеющимся данным, только очень крупными особями длиной 45 — 60 см (там нет ни мальков, ни сеголетков, ни созревающих рыб, а созревает ставрида в возрасте 2 — 3 лет). Таким образом, эти факты могут трактоваться лишь как доказательства существования временных псевдопопуляций (маловероятным представляется даже обособление зависимых популяций) , которые не только существуют за счет постоянного пополнения из прибрежного запаса, но и являются фактически его излишком. Выселение перуанской ставриды в океан вполне объяснимо ростом ее численности в неритической зоне. Так, согласно рыбопромысловой статистике ФАО (Продовольственная и сельскохозяйственная организация ООН), уловы этого вида в прибрежных водах Перу и Чили (они, видимо, отражают и истинную численность ставриды) увеличились со 100 тыс. т в 1970 г. и 300 — 400 тыс. т в 1974 — 1976 гг. до 1100 — 1300 тыс. т в 1978 — 1980 гг., а в открытом океане эта рыба была обнаружена в 1978 г.
Какой же биологический смысл могут иметь безвозвратные уходы прибрежно-пелагических рыб от родных берегов? Какую выгоду получает неритическая популяция, посылая на гибель в открытом океане большие группы «лишних» особей? Такая постановка вопросов представляется не совсем корректной (ибо отнюдь не все в живой природе имеет прямое приспособительное значение), но ответ на них все-таки существует: в результате таких выселений появляется потенциальная возможность расширить видовой ареал, включив в него новые неритические участки, лежащие за «барьером» открытоводных просторов.
То, что я написал здесь, еще два-три года назад показалось бы очень спорным большинству моих коллег-ихтиологов. Обнаружение ставриды в юго-восточной части Тихого океана далеко за пределами неритической зоны, в которой ей, казалось бы, полагалось пребывать, послужило основой для рассуждения об особых океанических популяциях этой рыбы, совершенно самостоятельных и не имеющих никакого отношения к прибрежным группировкам своего вида. Высказывались даже предположения о существовании своего рода «ставридного пояса», сплошным кольцом опоясывающего земной шар в районе «ревущих сороковых» широт южного полушария, и специально посланные суда искали там (да и в других участках открытого океана) «большую рыбу» — обособленные популяции массовых прибрежных видов. Мои представления (а я неоднократно излагал их в докладах на разных ученых собраниях и просто в разговорах с товарищами по профессии) не встречали тогда особой поддержки, хотя не удостаивались и критики. Сейчас, однако, оппонентов этим взглядам практически не осталось, и обо всей этой истории можно было бы и не упоминать, если б не одно важное обстоятельство, на которое хочется обратить специальное внимание: ведь если все так, как здесь написано, то рассчитывать на неограниченно долгое существование той же перуанско-чилийской ставриды в дальнем океане не приходится. Цикличность процесса неизбежно предполагает грядущий спад численности и даже почти полное исчезновение этой рыбы из открытых для международного рыболовства вод. Правда, в те же сроки в каких-то иных участках Мирового океана могут появиться псевдопопуляции других видов. Прогнозировать ход этих пульсационных процессов — одна из важнейших, на мой взгляд, задач прикладной рыбохозяйственной науки.