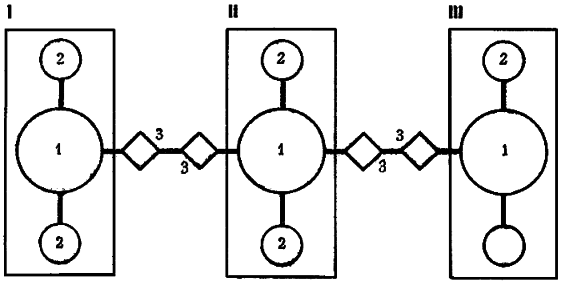
Рис. 17. Блок-схема федерации северного лесного муравья
I, II, III — колонии, входящие в состав федерации.
1 — материнские муравейники, 2 — отводки, 3 — буферные гнезда
Появление вторичных материнских муравейников симптоматично для федерации. Оно показывает, что даже очень крупные отводки, которые в состоянии сами образовывать дочерние муравейники, при интенсивных обменах удерживаются в составе колоний. В нашем примере муравейник 197 имеет диаметр купола 2,2 м, а его отводок 198 — 1,9 м. Обе величины предельны для данного вида. В сравнении с этими гнездами два других материнских муравейника выглядят карликами (диаметр купола гнезда 190 равен 1,2 м, гнезда 185 — 1,3 м). Тем не менее никакой иерархии колоний нет, так как при объединении между ними были сооружены буферные гнезда* — постоянные промежуточные почки. Буфером между колониями 1 и 2 служат муравейники 187-188, между колониями 2 и 3 — 182 и 199.
Объединение колоний, по-видимому, не обошлось без предварительных столкновений на территории. Об этом свидетельствуют имеющиеся в пограничных зонах колоний следы старых брошенных небольших гнезд, что обычно указывает на острую территориальную конкуренцию (см. рисунок). Однако теперь сражения за территорию в прошлом, все муравейники оказались связанными общей цепью обменов. Их интенсивность очень высока во всех звеньях федерации. Интенсивность переноски особей на основных дорогах, связывающих гнезда-доминанты, достигают 40 и белее в минуту (до 200). На остальных дорогах интенсивность — 15-30 переносов в минуту, что соответствует максимальным величинам в отдельных колониях. В колонии обмены в 40 и более переносов в минуту отмечены только в период формирования отводка.
Все обмены осуществляются через промежуточные почки.
Называть эти гнезда промежуточными почками можно только по аналогии. Фактически буферные гнезда — это особый тип летних гнезд, выполняющих «федеральные функции». Здесь имеется постоянное, частично смешанное население, среди которого высока доля муравьев-носильщиков. Благодаря этим муравейникам становится возможным эстафетный обмен между материнскими семьями. В обменных операциях участвуют десятки тысяч особей. Часть из них занята поддержанием в порядке буферных гнезд, другие осуществляют переносы.
Каким образом осуществляется запуск этого мощного механизма интеграции, неизвестно. С точки зрения экономики отдельных семей столь интенсивные обмены, по-видимому, мало рациональны. Но тем не менее это происходит, в виде федерации панмуравейник получает свое приближенное воплощение. Если в отдельной семье или колонии социальная регуляция колонн или входящих в колонию муравейников возникает как механизм, обеспечивающий общий рост населения, то в федерации направленность регуляции меняется. Здесь единственная функция обменных операций — ограничение общей численности муравьев в системе. Результатом действий регуляторных механизмов в семье и колонии является увеличение числа подсистем (колонн, семей), расселение вида; результатом регуляции в федерации — сохранение основных структурных единиц. О расселении речь не идет, поскольку федерации возникают в условиях, когда расселяться муравьям уже некуда.
И все это достигается одними и теми же средствами — обменами. Чем жестче территориальная конкуренция, тем интенсивнее обмены.
Данный принцип оправдан не только в федерациях. В эксперименте, проведенном на колонии муравьев-фаэтончиков в Туркмении в 1972 г., искусственное сокращение кормовых участков материнского и дочернего муравейников вызвало увеличение интенсивности обменов в 10-15 раз. Одновременно у подопытных муравейников появилась охраняемая территория, не свойственная фаэтончикам в обычных условиях.
Очень четкое разделение охраняемых территорий всех муравейников наблюдается в федерации. Такая взаимосвязь двух весьма различных по сути явлений понятна, так как оба они вытекают из объективной необходимости более эффективной регуляции в условиях жестких территориальных ограничений.
Сочетание четкого разграничения кормовых участков с интенсивными обменами обеспечивает стабильность федерации как системы муравейников. Устойчивость федерации значительно выше, чем обособленной колонии. Хотя разрыв отдельных связей между колониями возможен, тут же возникают новые, выполняющие ту же функцию. Сами же муравейники почти не меняются. За пять лет наблюдений в рассматриваемой федерации северного лесного муравья не погибло ни одного желтого муравейника и не возникло ни одного нового. Соотношение гнезд по размерам сохранилось приблизительно одинаковое, чего не бывает столь продолжительное время в самостоятельной колонии. Фактически неизменными по интенсивности остались и обмены, производимые муравьями летом.
Наименее устойчивыми элементами в федерации оказываются межколониальные обменные дороги и связанные с ними буферные гнезда. Они могут сдвигаться на местности, но сохраняют при этом полную функциональную преемственность. Таким образом в федерации реализуется стратегическая задача: обеспечение регулирования исключительно поведенческими средствами плотности поселения вида. Регуляция и устойчивость при максимальной для данных условий плотности поселения популяции. Такая задача в мире насекомых оказалась под силу только муравьям, точнее — лишь некоторым их видам. Все муравейники, объединенные в находящиеся под наблюдением федерации, активны, а гнезда — в хорошем состоянии. Почему? Ведь вся система функционирует в напряженном режиме (об этом можно судить хотя бы по интенсивности обменов). Может быть, следует сделать допущение, что именно такой режим и является лучшим средством, предохраняющим муравейники от старения и вырождения. Точнее, единственным шансом сохранить активное состояние в условиях перенаселения. Мы можем проверить это предположение, ознакомившись с развитием муравейников тех видов, у которых есть колонии, но не отмечены федерации. Например, волосистого лесного муравья. Гнезда у этого вида, как правило, крупнее гнезд северного лесного муравья, обычно обилие кормовых почек и отводков, но федерации нет. Собственно и не важно, может ли волосистый муравей создать федерацию в принципе. Достаточно взять какой-либо крупный комплекс, в котором федераций нет, а плотность поселения угрожающе высока. Ведь всякая возможность реализуется не всегда. В частности, старый муравейник, стабилизировавшийся в своем одиночном существовании, не дает отводков и не войдет в федерацию. Даже если это будет муравейник столь склонного к федерациям северного лесного муравья.
Весьма обычен комплекс муравейников, «законсервировавшихся» раньше, чем сложились условия перенаселения, подталкивающие муравьев к объединению в федерации. Но за счет отдельных, разбросанных по комплексу колоний плотность поселения все-таки возрастает и достигает критического уровня. Территориальная конкуренция обостряется, разыгрываются ожесточенные сражения между соседними муравейниками, но за этими сражениями не следует объединение. Периодические сражения — плохой регулятор численности. Вскоре из-за нехватки белковой пищи изменяется пищевой баланс семей (возрастает доля пади), сокращается количество выкармливаемой молоди. Теперь уже не до отводков.
Семьи становятся все нетерпимее к чужим особям, в частности к чужим самкам. Замена старых самок молодыми затрудняется. Семьи приходят в упадок, погибают. От комплекса остаются отдельные, едва влачащие существование муравейники или периферийное кольцо муравейников, из которых со временем начнется повторное заселение центральной части комплекса.
Значит, одной системы охраняемых территорий еще недостаточно, чтобы поддерживать комплекс в активном состоянии долгое время. В комплексе остается стихийное начало, ведущее к подъемам и депрессиям, подобным зигзагам движения численности у одиночных насекомых. Конечно, амплитуды колебаний здесь не столь значительны, да и катастрофические спады случаются очень редко. Но все же время от времени они происходят.