Преобразование энергии в живой клетке описывается следующей общей схемой:
Энергетические ресурсы → ΔμI → работа
где ΔμI – трансмембранная разность электрохимических потенциалов иона I. Следовательно, процессы утилизации энергии и совершения за счет нее работы оказываются сопряжены через образование и использование ΔμI. Поэтому данный ион может быть назван сопрягающим ионом. Основным сопрягающим ионом в клетке эукариот является Н+, и соответственно ΔμН+ является основной конвертируемой формой запасания энергии. Вторым по значимости сопрягающим ионом является Na+ (ΔμNa+). В то время как Сa2+, K+ и Cl- не используются для совершения какой-либо работы.
Биологическое окисление – это процесс дегидрирования субстрата с помощью промежуточных переносчиков водорода и его конечного акцептора. Если в роли конечного акцептора выступает кислород, процесс называется аэробным окислением или тканевым дыханием, если конечный акцептор представлен не кислородом – анаэробным окислением. Анаэробное окисление имеет ограниченное значение в организме человека. Основная функция биологического окисления – обеспечение клетки энергией в доступной форме.
Тканевое дыхание – процесс окисления водорода кислородом до воды ферментами цепи тканевого дыхания. Оно протекает по следующей схеме:
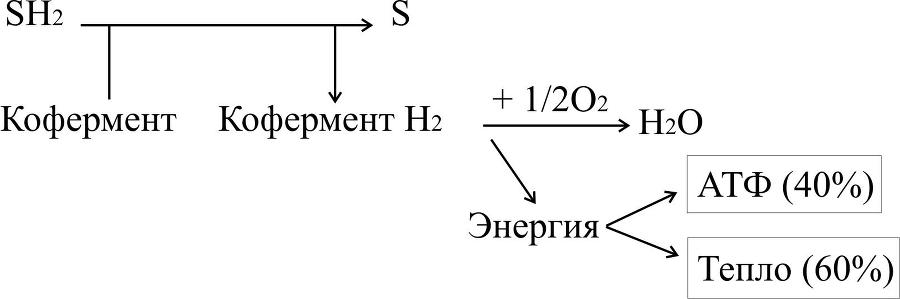
Вещество окисляется, если отдает электроны или одновременно электроны и протоны (атомы водорода), или присоединяет кислород. Способность молекулы отдавать электроны другой молекуле определяется окислительно-восстановительным потенциалом (редокс-потенциалом). Любое соединение может отдавать электроны только веществу с более высоким окислительно-восстановительным потенциалом. Окислитель и восстановитель всегда образуют сопряженную пару.
Выделяют 2 типа окисляемых субстратов:
1. Пиридинзависимые – спиртовые или альдегидные – изоцитрат, α-кетоглутарат, пируват, малат, глутамат, β-гидроксиацил-КоА, β-гидроксибутират, – в их дегидрировании участвуют НАД-зависимые дегидрогеназы.
2. флавинзависимые – являются производными углеводородов – сукцинат, ацил-КоА, глицерол-3-фосфат, холин – при дегидрировании передают водород на ФАД-зависимые дегидрогеназы.
Цепь тканевого дыхания – последовательность переносчиков протонов водорода (Н+) и электронов от окисляемого субстрата на кислород, локализованных на внутренней мембране митохондрий.

Рис. 10.1. Схема ЦТД
Компоненты ЦТД:
1. НАД-зависимые дегидрогеназы дегидрируют пиридинзависимые субстраты и акцептируют 2ē и один Н+.
2. ФАД (ФМН) - зависимые дегидрогеназы акцептируют 2 атома водорода (2Н+ и 2ē). ФМН – зависимая дегидрогеназа дегидрирует только НАДН, в то время как ФАД-дегидрогеназы окисляют флавинзависимые субстраты.
3. Жирорастворимый переносчик убихинон (кофермент Q, КоQ) – свободно перемещается по мембране митохондрий и акцептирует два атома водорода и превращается в КоQH2 (восстановленная форма – убихинол).
4. Система цитохромов – переносит только электроны. Цитохромы железосодержащие белки, простетическая группа которых по структуре напоминает гем. В отличие от гема атом железа в цитохроме может обратимо переходить из двух – в трехвалентное состояние (Fe3+ + ē → Fe2+). Это и обеспечивает участие цитохрома в транспорте электронов. Цитохромы действуют в порядке возрастания их редокс-потенциала и в дыхательной цепи располагаются следующим образом: b-с1-с-а-а3. Два последних работают в ассоциации как один фермент цитохромоксидаза аа3. Цитохромоксидаза состоит из 6 субъединиц (2 - цитохрома а и 4 - цитохрома а3). В цитохроме а3 кроме железа имеются атомы меди и он передает электроны непосредственно на кислород. Атом кислорода при этом заряжается отрицательно и приобретает способность взаимодействовать с протонами с образованием метаболической воды.
Железосерные белки (FeS) – содержат негемовое железо и участвуют в окислительно-восстановительных процессах, протекающих по одноэлектронному механизму и ассоциированы с флавопротеинами и цитохромом b.
Структурная организация цепи тканевого дыхания
Компоненты дыхательной цепи во внутренней мембране михохондрий формируют комплексы:
1. I комплекс (НАДН-КоQН2-редуктаза) – принимает электороны от митохондриального НАДН и транспортирует их на КоQ. Протоны транспортируются в межмембранное пространство. Промежуточным акцептором и переносчиком протонов и электронов являются ФМН и железосерные белки. I комплекс разделяет поток электронов и протонов.
2. II комплекс – сукцинат – КоQ - редуктаза – включает ФАД- зависимые дегидрогеназы и железосерные белки. Он транспортирует электроны и протоны от флавинзависимых субстратов на убихинон, с образованием промежуточного ФАДН2.
Убихинон легко перемещается по мембране и передает электроны на III комплекс.
3. III комплекс – КоQН2 - цитохром с - редуктаза – имеет в своем составе цитохромы b и с1, а также железосерные белки. Функционирование КоQ с III комплексом приводит к разделению потока протонов и электронов: протоны из матрикса перекачиваются в межмембранное пространство митохондрий, а электроны транспортируются далее по ЦТД.
4. IV комплекс – цитохром а - цитохромоксидаза – содержит цитохромоксидазу и транспортирует электроны на кислород с промежуточного переносчика цитохрома с, который является подвижным компонентом цепи.
Существует 2 разновидности ЦТД:
1. Полная цепь – в нее вступают пиридинзависимые субстраты и предают атомы водорода на НАД-зависимые дегидрогеназы
2. Неполная (укороченная или редуцированная) ЦТД в которой атомы водорода передаются от ФАД-зависимых субстратов, в обход первого комплекса.
Окислительное фосфорилирование АТФ
Окислительное фосфорилирование – процесс образования АТФ, сопряженный с транспортом электронов по цепи тканевого дыхания от окисляемого субстрата на кислород. Электроны всегда стремятся переходить от электроотрицательных систем к электроположительным, поэтому их транспорт по ЦТД сопровождается снижением свободной энергии. В дыхательной цепи на каждом этапе снижение свободной энергии происходит ступенчато. При этом можно выделить три участка, в которых перенос электронов сопровождается относительно большим снижением свободной энергии. Эти этапы способны обеспечить энергией синтез АТФ, так как количество выделяющейся свободной энергии приблизительно равно энергии, необходимой для синтеза АТФ из АДФ и фосфата.
Для объяснения механизмов сопряжения дыхания и фосфорилирования выдвинут ряд гипотез.
Механохимическая или конформационная (Грин-Бойера).
В процессе переноса протонов и электронов изменяется конформация белков-ферментов. Они переходят в новое, богатое энергией конформационное состояние, а затем при возвращении в исходную конформацию отдают энергию для синтеза АТФ.
Гипотеза химического сопряжения (Липмана).
В сопряжении дыхания и фосфорилирования участвуют «сопрягающие» вещества. Они акцептируют протоны и электроны и взаимодействуют с Н3РО4. В момент отдачи протонов и электронов связь с фосфатом становится макроэргической и фосфатная группа передается на АДФ с образованием АТФ путем субстратного фосфорилирования. Гипотеза логична, однако до сих пор не выделены «сопрягающие» вещества.