Итак, иллюзия. Но мы-то знаем, что любая иллюзия есть отражение нормальной работы зрительного аппарата, нарочно поставленного в непривычные условия.
Так и в опытах Бертулиса и Майкла. Эксперимент удается лишь при равенстве пространственных частот цветной и черно-белой решеток. В противном случае эффекта нет. Цветоощущающие поля коры, настроенные на выделение цветных решеток, работают, по-видимому, так же, как поля лягушачьей НКТ. Если в первой фазе эксперимента они возбуждены и дают сигнал «Цвет есть!», то во второй фазе белый луч выглядит для них уменьшением яркости «их» цвета (ведь белый свет потому и белый, что не имеет резкого преобладания какого-то одного тона). То есть получается «скольжение» по красно-зеленой или сине-желтой оси сигналов. И вместо красного видится зеленый тон, вместо синего – желтый. Все те же «качели»...
Отсюда Глезер и его коллеги по Лаборатории заключают: цветовое зрение формируется благодаря тому, что в затылочной коре, помимо черно-белых полей, осуществляющих кусочное квазиголографическое представление изображений, обязаны быть нейроны, таким же квазиголографическим способом отражающие окраску. Это и есть те самые каналы передачи цвета, существование которых давно уже предполагалось.
Альгис Бертулис обнаружил в 1980...1982 гг. очень интересную их особенность: каналы (то есть составляющие их рецептивные поля) способны передавать только сравнительно низкие пространственные частоты. Это значит – не выше 10 цикл/град, если краски отстоят друг от друга далеко по спектру (скажем, красный и фиолетовый), и всего 2...3 цикл/град, если надо распознавать красный и оранжевый, зеленый и голубой, синий и фиолетовый.
Казалось бы, при таких условиях мы принципиально не в состоянии различать мелкие детали цветных изображений. Но опыты говорят иное. При нормальном освещении четкость цветового зрения около одной угловой минуты, то есть в худшем случае шестикратно, а в лучшем – тридцатикратно выше, чем следует из опытов Бертулиса. Ошибка? Некорректно поставленный эксперимент? Нет, работа безупречна. Просто цветовому зрению помогает черно-белое! Как? Чтобы рассказать об этом, придется вспомнить о некоторых опытах Ярбуса.
Глаза наши все время находятся в движении, но что случится, если их остановить? Для этого Ярбус изобрел в начале 60-х гг. присоску (мы ее уже упоминали) – крошечный проекционный аппаратик, куда можно вставлять картинки, тест-объекты. Аппаратик столь миниатюрен, что сила атмосферного давления «приклеивает» его прямо к глазному яблоку, так что тест-объект оказывается совершенно неподвижным относительно сетчатки. И... спустя одну-две секунды изображение исчезает! Вместо картинки в поле зрения возникает светло-серая пелена, которую можно увидеть, закрыв глаза или попав в темную комнату. Пелена, как мы знаем, – результат спонтанной активности ганглиозных клеток сетчатки.
Куда же девалось изображение? А никуда. Легкий удар кончиком карандаша по тест-объекту – и он вновь возникает, чтобы через секунду опять пропасть. Вот теперь все ясно: удар нарушил неподвижность картинки относительно сетчатки. Выходит, только движение (глаза или картинки, неважно) порождает зрительный образ. Оно принципиально необходимо, чтобы зрение работало. И действительно, стоит ввести перед тест-объектом что-нибудь движущееся, как этот предмет оказывается прекрасно различим на фоне серой пелены – «нуль-цвета», как назвал его Ярбус.
Совершенно неожиданное следствие вытекало из опытов: сам по себе свет еще не обеспечивает видения.
Зрительная система равно отображает «нуль-цветом» и полную темноту, и неподвижность изображения относительно сетчатки. Более того, пусть яркость этого неподвижного тест-объекта сколь угодно велика – глаз этого не заметит. «Даже раскаленная, слепяще яркая нить электрической лампочки становится невидимой», – писал Ярбус. Какой же элемент выключается? Скорее всего, сетчатка. Ведь второй глаз, на котором нет присоски, продолжает все великолепно различать. Значит, после хиазмы все структуры зрительного аппарата, через которые проходят сигналы от обеих сетчаток, действуют нормально. А в глазу с присоской картинка попадает все время на одни и те же фоторецепторы, они воспринимают постоянную, никак не изменяющуюся яркость, хотя нуждаются в ином: чтобы световое воздействие на них было все время разным. Для этого глаза и движутся.
Что случится, если «нуль-цвет» появится на фоне какого-нибудь видимого изображения? Ярбус изготовил присоску, тест-объект которой закрывал лишь часть поля зрения. Испытуемый увидел странную вещь: тест-объект (это была просто белая бумажка) превратился в какого-то хамелеона. Стоило направить взор на зеленый щит, и она становилась зеленой, на фоне красного – красной, в несколько секунд полностью перекрашиваясь и совершенно сливаясь с ним. Бумажку заменили цветной, но это, как и следовало ожидать, совершенно не повлияло на ее «перекрасочные» свойства.
Возникло противоречие. С одной стороны, неподвижный тест-объект обязан вызвать «нуль-цвет» на том участке изображения, куда он проецируется, попадая на сетчатку. С другой стороны, зрительный тракт с этим не желает считаться и подменяет «нуль-цвет» другим, зависящим от цвета изображения. Значит, фоторецепторы фоторецепторами, а в высших структурах зрительной системы работает некий маляр.
Как он выглядит нейрофизиологически?
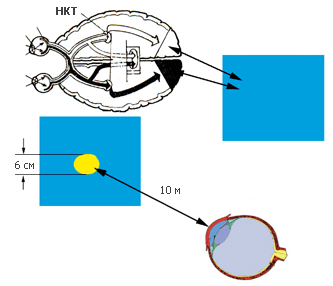
Рис. 57. Опыты каунасского исследователя Альгиса Бертулиса, ученика В.Д. Глезера, продемонстрировали, что более яркий фон «закрашивает» детали малого размера, и на большом расстоянии мы их просто не видим! Например, белое пятно диаметром 6 см сливается с фоном (в данном случае голубым) уже на расстоянии 10 м
Опыты Бертулиса и его коллег продемонстрировали, что эффект «прокрашивания» можно получить без присоски, во время нормального рассматривания. Нужно лишь установить яркость объекта и фона одинаковыми, а сам объект сделать не слишком большим, примерно в 20 угловых минут (так выглядит с десятиметрового расстояния кружок диаметром шесть сантиметров). Разница в цвете объекта и фона в этом случае может быть огромной, и все-таки объект будет окрашен в цвет фона, ассимилирован.
Выходит, сами по себе цветовые каналы еще не способны дать сведения о цвете малых объектов. Чтобы цветоощущение заработало, надо увеличить яркость объекта, повысить его контраст с фоном. На яркость же реагируют черно-белые, ахроматические поля. Как только они выделили предмет, отличили его от фона, вступают в игру цветочувствительные поля и присваивают предмету цветовой тон – «прокрашивают».
Ведь различия в яркости – это пример простейшей, вырожденной текстуры. А для улавливания текстурных различий имеется ахроматический канал, он благодаря своим небольшим рецептивным полям (нейронов затылочной коры) обладает гораздо лучшей разрешающей способностью, нежели цветоощущающие каналы, то есть воспринимает мелкие и мельчайшие детали.
Это очень хорошо видно, когда на ассимилированный по цвету кружочек накладывают решетку, а на фон – точно такую же, но по-иному ориентированную. Цвет кружочка сразу проявляется, хотя его яркость осталась прежней («невидимой») относительно фона. Ведь текстурный канал уже выделил форму объекта, так что цветовому не составляет труда окрасить выделенное.
Итак, окончательное суждение о цвете зависит и от яркости предмета относительно фона, и от текстуры. Это объясняет множество эффектов, известных каждому из собственного опыта.
Вот на красном полотнище нарисована зеленая ветвь – и ощущение зеленого цвета оказывается связанным и с этим красным цветом, и с цветом неба, на фоне которого смотрится полотнище, и с общей яркостью освещения. Каждый новый цветовой тон, лежащий на фоне иной краски, выглядит очередной фигуркой удивительной цветовой «матрешки»: окраска каждой внутренней фигуры зависит от окраски внешнего по отношению к ней фона. Так что фон способен и «поднять», и «убить» положенный на него цвет, как прекрасно чувствуют не только художники, но и многие дамы, следящие за своими нарядами. Все дело в сложной игре сигналов-чисел, поступающих в высшие отделы мозга от нейронов затылочной коры.