Но вернемся к теории Юнга-Гельмгольца. Она неплохо объясняет, как из цветов спектра образуются различные краски. Она подсказывает, каким способом можно «обмануть» глаз и показать ему один и тот же цвет, смешивая пары совершенно различных лучей. Для этого нужно только соответствующим образом возбудить различные колбочки. Существует множество комбинаций лучей, воспринимаемых глазом как белый свет: его дадут такие пары, как, например, имеющие длины волн 486 и 590 нанометров (голубой и оранжевый), 467 и 572 нанометра (синий и желто-зеленый), 494 и 640 нанометров (красный и зеленый), и так далее, и так далее... Вместе с тем красный и зеленый лучи могут дать великолепный желтый тон, который, кроме того, легко составить из оранжевого и зеленовато-синего света... Рецептов создания любого цвета, лежащего в средней части спектра, оказывается тысячи. Обо всем этом убедительно говорят учебники. Умалчивают они лишь о том, чего теория не объясняет.
А трехкомпоненная теория плохо объясняет некоторые расстройства зрения. Например, почему некоторые дальтоники видят только синие лучи, а всё остальное – в черно-белом варианте. Ведь белое по этой теории есть результат сочетания трех сигналов от трех типов колбочек, и если это так, должны быть ощущения других цветов. Словом, когда нейрофизиологи смогли подключить к ганглиозным клеткам сетчатки свои приборы и стали освещать ее не белым светом, а разноцветными лучами, оказалось, что сигналы от колбочек есть, только они сочетаются между собой совсем не так, как мыслилось по теории Юнга-Гельмгольца. Что поделать, наука на месте не стоит, и у любой теории есть вершина и спад...
Основываясь на феномене «сине-белых» дальтонических расстройств, известный немецкий физиолог Эвальд Геринг выдвинул в 1874 г. гипотезу, весьма расходившуюся с господствовавшей тогда трехкомпонентной: вместо сложения сигналов основой было вычитание. Геринг утверждал, что в чувствительных элементах глаза находятся три вещества, из которых одно распадается под действием красных лучей и восстанавливается от зеленых, другое претерпевает такие же изменения благодаря синим и желтым лучам, а третье чувствительно к черным и белым. Это казалось нелепостью: вы когда-нибудь видели черный свет? Да к тому же никаких веществ такого рода найти не удалось, а авторитет Гельмгольца, и вполне заслуженно, был высок.
Словом, о гипотезе Геринга вспоминали в учебниках не более как об историческом факте, чуть ли не курьезе. Но судьба почему-то любит неудачников с острым умом. Девяносто лет спустя после публикации работы Геринга вышла из печати статья Роберта Де Валуа и Джорджа Джекобса: ганглиозные клетки сетчатки глаза лягушки работают «по Герингу»!
Помните, мы говорили об обратных связях в сетчатке? Мы увидели там систему, благодаря которой в мозг поступают из ганглиозных клеток сигналы, свидетельствующие не о яркости света на данном участке, а только об отклонениях этой яркости от некоего среднего значения, средней освещенности. Вверх – белый свет, а вниз... иначе как черным его не назовешь!
Такое же положение и с цветовыми сигналами. Цветовые лучи воспринимаются лягушачьей сетчаткой с помощью полей ганглиозных клеток. Но поля эти по своим ответам гораздо сложнее, чем черно-белые. До тех пор, пока никакого света на него не подано, ганглиозная клетка отправляет в мозг сигналы спонтанной активности – редкие, как бы случайные импульсы. Благодаря им даже в полной темноте мы видим не черноту перед глазами, а как бы колеблющуюся серую пелену.
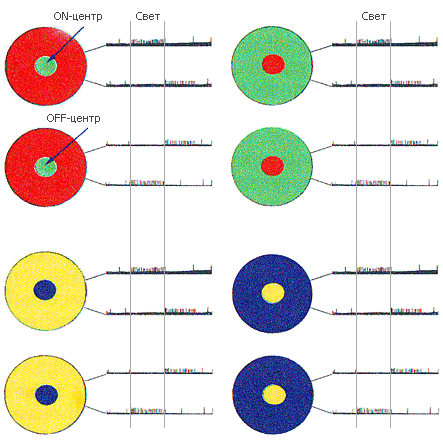
Рис. 56. Цветовые поля наружного коленчатого тела (НКТ между сетчаткой и зрительной корой), «взвешивающие» пары цветов: какого цвета больше? Всего возможно восемь комбинаций: четыре комбинации с ON-центром сложного поля и четыре комбинации с OFF-центром сложного поля. Этим обеспечиваются ответы о перепады яркостей «свет – темнота» и «темнота – свет». Всё это происходит задолго до работы зрительных полей коры
Но вот исследователь начинает проверять поле с зеленым «он»-центром и красной «офф»-периферией (левый верхний рисунок). Дает зеленый свет на центр – спонтанная активность сменяется дробью импульсов: «Есть свет!». Выключает свет – сигналов нет, затормаживается на некоторое время даже спонтанная активность.
С периферией, как и положено, зависимость обратная. Красное световое кольцо угашает спонтанную активность, а выключение света заставляет ганглиозную клетку дать сигнал.
У других трех типов «красно-зеленых» ганглиозных клеток поля имеют либо зеленый «офф»-центр и красную «он»-периферию, либо красный «офф»-центр и зеленую «он»-периферию, либо, наконец, красный «он»-центр и зеленую «офф»-периферию... Уфф! И еще четыре такие же пары существуют для желтых и синих лучей. (У приматов такой работой заняты нейроны коры.)
Нейроны же НКТ лягушачьего мозга предстают в роли «весов», взвешивающих цветовые сигналы. Первый тип возбуждается от красного и тормозится от зеленого, второй тип от красного тормозится, а от зеленого возбуждается. Третий и четвертый типы таким же манером обращаются с голубым и желтым цветами.
Де Валуа обнаружил, что реакция таких клеток НКТ зависит не только от длины волны света, падающего на сетчатку. Клетки отвечают на направление сдвига длины этой волны. Положим, мы исследуем клетку, тормозящуюся от красного и возбуждающуюся от зеленого света. Но сменив освещающий красный луч на желтый, мы все равно получим возбуждение: ведь произошел сдвиг от более длинноволнового, красного света, к более коротковолновому, желтому. Зато если возбуждающий зеленый сменится тем же желтым (сдвиг от коротких волн к длинным), возникнет тормозная реакция.
То есть ответ ганглиозной клетки зависит не от абсолютного значения длины волн, а от длины волны «предыдущего» света. По трехкомпонентной теории такого просто быть не может.
И когда изменяется яркость возбуждающего зеленого света – клетка НКТ отслеживает своим сигналом эти изменения.
Аналогично действуют и другие клетки НКТ такого рода.
Смотрите, какая удивительная получается картина: нейронам безразлично, какова причина «позеленения» или «покраснения» цвета, падающего на поле данной клетки НКТ. Изменилась ли яркость (вышло солнце из-за туч или спряталось), стало ли иным сочетание лучей разных длин волн в спектре (в зените солнце или оно у горизонта со своей алой зарей), – в любом случае реакция нейронов НКТ будет соответствовать только направлению этого изменения. Словно качели в детском саду: Сережа поднимается вверх – Петя опускается...
Однако из «качелей» следует и другой, куда более важный вывод: с помощью рецепторов сетчатки и нейронов НКТ лягушка не в состоянии увидеть цвет. И мы тоже. Самое большее – это отметим, в каком направлении по спектру (от красного конца к фиолетовому или наоборот) изменяется окраска того участка изображения, который попал на поле данного нейрона. Значит, и тут встречаемся все с тем же принципом: цвет «конструируется» в высших отделах мозга, а сетчатка только поставляет для этого «строительные материалы».
Действительно, в 1977 г. ученик Глезера, каунасский физиолог Альгис Бертулис со своими коллегами (а год спустя американец Д. Майкл) обнаружили в затылочной коре обезьяны поля – такие же, какие для черно-белых стимулов нашли у кошек Хьюбел и Визел. Эти новые поля отвечают на всевозможно ориентированные полоски вполне определенного цвета. Оставалось только доказать, что нейроны коры реагируют на цветные решетки – красно-зеленые и желто-синие.
Прямые опыты с нейронами и косвенные – психологические – дали одинаковые по смыслу результаты. Например, испытуемому показывают две решетки: вертикальную красно-черную и горизонтальную сине-черную. Затем появляются вертикальная и горизонтальная черно-белые решетки тех же пространственных частот. Но испытуемый видит... черно-зеленую и черно-желтую! Почему? Потому что по закону последовательного цветового контраста «иллюзорный» цвет является дополнительным к истинному, предыдущему, соответственно работе нейронных «качелей».