Исследования, проводимые по селекции мышей на различный размер мозга, также показали, что генотипические факторы оказывают влияние на способность к экстраполяции у животных разных линий. Селекция на большой и малый вес мозга была начата на основе гетерогенной популяции, полученной путем скрещивания шести инбредных линий лабораторных мышей. Критерием для селекции служила регрессия, связывающая два признака — все тела и вес мозга. Уже с четвертого поколения установилось достоверное (р<0,001) различие среднего веса мозга у сублиний «большой» и «малый мозг». К 14-му поколению это различие достигло 63 мг. Доля правильных решений у сублинии «малый мозг» оказалась достоверно выше 50 %-ного случайного уровня, но начиная с 13-го поколения селекции правильные решения у обеих сублиний снизились до 50 %-ного случайного уровня. На основании проведенных исследований может быть сделан вывод, что различие в весе мозга не является ведущим фактором у особей, принадлежащих к одному виду, в решении элементарной логической задачи. Однако полученные данные указывают на изменение способности к экстраполяции у мышей селекционированных линий.
Проведенные опыты на крысах, мышах и сравнение способности к экстраполяции красных лисиц (Vulpes vulpes) и серебристо-черных (Vulpes fulvus) показали влияние генотипа на элементарную рассудочную деятельность у животных (Сотская, 1974). Сравнение уровня рассудочной деятельности диких и доместицированных форм показало, что доместицированные животные, как правил, обладают менее развитой рассудочной деятельностью, чем их дикие предки (Крушинский, 1977). Наиболее вероятной причиной такого различия является ослабление пресса естественного отбора у доместицированных животных. Ослабление действия естественного отбора приводит к увеличению изменчивости фенотипа и распаду полигенных систем, обеспечивающих наиболее высокие формы приспособления организмов к многообразным изменениям среды обитания. Такая дезинтеграция коадаптированных генетических систем является результатом дестабилизирующего отбора (Беляев, 1972).
При исследовании рассудочной деятельности у животных, которое проводится разными методами, мы столкнулись с неожиданным фактом появлением при решении предъявляемых тестов неадекватных форм поведения. Практически среди животных всех исследованных таксономических групп встречаются особи, у которых после нескольких правильных решений поведение во время эксперимента настолько меняется, что дает основание предполагать развитие какой-то патологии:
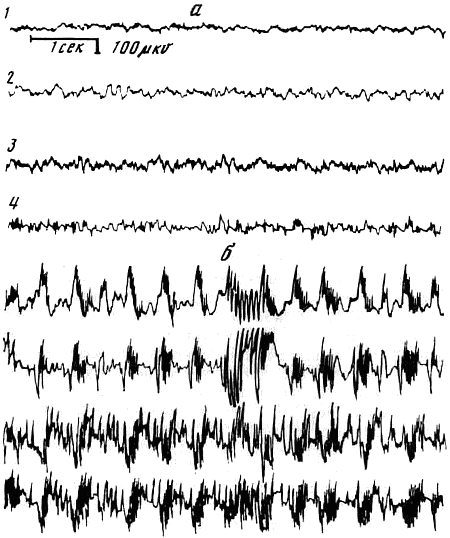
а — фон; б — после решений задачи; 1 — сенсомоторная область коры; 2 — дорсальный гиппокамп, 3 — септум; 4 — латеральное поле гипоталамуса
Электрофизиологические исследования, проводимые на крысах и черепахах, показали, что при решении предъявляемых задач наблюдаются высоковольтные разряды и комплексы пик-волна, которые характерны для эпилептического припадка. Патологические разряды регистрировались одновременно как в корковых, так и подкорковых структурах мозга (Семиохина и др., 1976) (рис. 4).
Результаты этих исследований позволяют прийти к заключению, что при решении незнакомых для животных логических задач может развиваться резкое возбуждение мозга, которое широко иррадиирует по отдельным мозговым структурам. Оно приводит к появлению неадекватных реакций поведения: животное начинает бояться экспериментальной обстановки, убегает от нее и т. д. Очевидно, можно предположить, что не только у животных, но и у людей напряжение рассудочной деятельности, которое неизбежно в период творческой работы, может привести к резкому возбуждению мозга и появлению ряда патологических симптомов.

Удивительная, но вполне объяснимая связь между рассудочной деятельностью и патологией выступает при селекции. В лаборатории постнатального онтогенеза ИБР АН СССР у двух доместицированных линий пасюков (Rattus norvegicus) Н. Л. Крушинской и И. Л. Дмитриевой ведется селекция на способность крыс к экстраполяции. Отбор проведен в четырех поколениях (см. табл. 2).
Никакого эффекта улучшения рассудочной деятельности не обнаружено. Наоборот, было установлено, что от поколения к поколению уменьшается число хорошо решающих крыс и увеличивается число особей, с которыми трудно проводить опыты. Доля правильных решений резко уменьшается после первого предъявления задачи. Это обусловлено большой «нервозностью» крыс, которые плохо адаптируются к экспериментальной обстановке: стремятся убежать из нее или забиваются в угол камеры. Подобное же явление наблюдалось ранее и в нашей лаборатории.
Полученные данные о появлении невротиков в линиях крыс, селекционированных по признаку высокой рассудочной деятельности, едва ли случайны. Наиболее логичное объяснение этого факта сводится к следующему. При селекции на высокий уровень рассудочной деятельности отбираются в первую очередь наиболее возбудимые особи. Такие животные весьма предрасположены к всевозможным формам неврозов. Развитие неврозов мешает выявлению рассудочной деятельности, что и выражается в уменьшении процента в селекционируемых линиях крыс, хорошо решающих задачи. Сказанное предположение может быть проверено экспериментально. Если оно справедливо, то введение селекционированным крысам седативных средств должно уменьшить уровень возбудимости и ослабить или полностью купировать неадекватные формы их поведения.
Одним из ярких выражений неадекватности поведения явилась парадоксальная реакция на отодвигающийся за ширмой корм в опытах с экстраполяцией. В этом случае крысы шли не за кормом, а в противоположном направлении. Введение таким крысам мепробамата полностью купировало эту парадоксальную форму поведения. Ряд крыс начал адекватно решать предъявленную задачу.
Приведенные данные являются убедительной иллюстрацией значения уровня возбудимости как модификатора поведения и согласуются с высказанной рабочей гипотезой.
Таким образом, поднятая проблема о значении центральных механизмов нервной деятельности в фенотипическом выражении отдельных, частных, генотипически обусловленных особенностей переросла в более общую проблему — о регуляторном значении центральных механизмов нервной системы в фенотипической гетерогении поведения. В основе этой гетерогении лежит взаимодействие основных нервных процессов возбуждения и торможения, которое в каждый текущий момент определяет состояние возбудимости нервной системы.
Фенотипическая гетерогения поведения может быть разделена на две принципиально разные группы. К первой следует отнести гетерогению, являющуюся отражением взаимодействия основных нервных процессов: возбуждения и торможения. Основной характеристикой этого типа гетерогении являются парабиотические стадии, которые проявляются при разном уровне возбудимости нервной системы. В этом случае защитные механизмы, приводящие к стабилизации системы, являются источником гетерогении поведения. Ко второй группе мы относим гетерогению, обусловленную тем, что разный уровень возбудимости нервной системы оказывает модифицирующее влияние на проявление и выражение специфических особенностей поведения.
Проведенное исследование показало огромную роль центральных физиологических механизмов в фенотипической изменчивости как нормальных поведенческих актов, так и патологических реакций организма. Вместе с тем оно дало возможность убедиться в том, что в основе фенотипической корреляции между нормальными и патологическими признаками поведения лежит универсальный центральный механизм нервной системы, интегральное выражение которого определяется уровнем ее возбудимости.
Вероятно, одним из наиболее актуальных направлений регуляции фенотипической изменчивости поведения как в норме, так и при патологии является управление центральными физическими механизмами функций нервной системы; управление уровнем возбудимости целого мозга.