Продолжая увеличивать промежуток времени между моментом приложения тормозного раздражителя к одному пункту и моментом испытания рефлекса с другого пункта, можно определить и скорость концентрирования угасательного торможения в кожном анализаторе.
Абсолютные величины времени концентрации тормозного процесса, как и времени его иррадиации, сильно зависят от индивидуальных особенностей подопытных животных, однако их соотношение оказалось довольно постоянным у всех испытуемых собак. Как правило, иррадиирование происходит в 4–5 раз быстрее, чем последующее концентрирование.
Иррадиация возбуждения. Опыт, показывающий иррадиацию возбудительного процесса, в некоторых отношениях напоминает описанные выше опыты с иррадиацией торможения.
У собаки вдоль задней ноги от плюсны до таза на примерно одинаковом расстоянии друг от друга приклеивали пять касалок (рис. 36). На действие самой нижней касалки (№ 1) вырабатывали условный рефлекс слюноотделения, подкрепляемый вливанием раствора кислоты в рот собаки. При первом испытании и другие касалки (№ 2, 3, 4 и 5) как сходные раздражители тоже вызывали слюноотделение. Поэтому необходимо было выработать дифференцирование этих касалок, применяя многократно касалку № 1 с подкреплением, а остальные касалки — без подкрепления. Теперь только касалка № 1 вызывала слюноотделение, а остальные превратились в тормозные сигналы.
После такой подготовки приступали к главной части опыта. Включали положительную касалку № 1 на 15 с и сразу после ее выключения действовали отдифференцированной касалкой № 2. Однако ее действие тоже вызывало слюноотделение. Это означало, что пункт кожного анализатора, соответствующий касалке № 2, обычно находящийся в тормозном состоянии, сразу после возникновения очага возбуждения в пункте, соответствующем касалке № 1, тоже оказался в возбужденном состоянии. Иначе говоря, возбуждение из пункта касалки № 1 в это время распространилось на пункт касалки № 2. Если испытать таким образом и какой-либо другой, более удаленный пункт кожного анализатора, то можно судить о районе такой иррадиации. Эти опыты показали, что иррадиирующее возбуждение по мере удаления от очага своего развития постепенно ослабевает.

Рис. 36. Опыт с иррадиацией возбуждения по корковым клеткам кожного анализатора:
1 — положительный условный раздражитель, 2, 3, 4, 5 — дифференцировочные раздражители
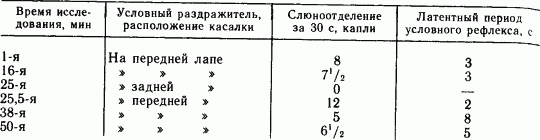
В другом варианте опытов под наблюдение была взята более обширная область кожного анализатора. Касалки располагали от конца передней лапы через все туловище до конца задней лапы. Первая касалка на конце передней лапы была положительным сигналом, а все остальные — дифференцировочными. Испытания дифференцировочными касалками показали, что уже через 0,5–1 с по прекращении положительного сигнала возбудительный процесс обнаруживается на соседних пунктах.
Таким образом, иррадиация возбуждения в коре мозга происходит гораздо быстрее, чем иррадиация торможения, и требует менее секунды для распространения по области кожного анализатора.
Через некоторое время после положительного сигнала соседние пункты анализатора вновь оказываются в прежнем тормозном состоянии. Это означает, что волна возбуждения успела уже разлиться по коре и вновь сосредоточиться в исходном пункте.
Движение основных процессов высшей нервной деятельности определяется не только свойствами иррадиации и концентрации, но и свойствами их взаимной индукции. Индукцией называется свойство каждого из основных нервных процессов вызывать вокруг себя и после себя противоположный процесс.
Положительная индукция из очагов угасательного и дифференцировочного торможения. В опытах с иррадиацией угасательного торможения по кожному анализатору часто наблюдают такое явление: повторение раздражения какой-либо одной касалкой без подкрепления способствует полному угасанию условного рефлекса с раздражаемого пункта кожи. Однако сразу после этого раздражение касалкой с другого отдаленного пункта выявляет этот рефлекс в резко усиленном виде. Возникший в первом пункте очаг сильного торможения индуцировал в отдаленном пункте возбуждение, привел этот пункт в состояние повышенной возбудимости.
Еще более четко явление положительной индукции было выявлено в специальных опытах на примере индукции, развиваемой дифференцировочным торможением. Так, у собаки вырабатывали условный пищевой рефлекс слюноотделения, в котором сигналом служило раздражение касалкой кожи передней лапы. Другая касалка была установлена на задней ноге. Ее применяли без подкреплений, так что скоро она приобрела тормозное значение дифференцировочного раздражителя. На включение дифференцировочной касалки слюноотделения не наступало, однако испробованный сразу после нее положительный раздражитель давал резко усиленный рефлекс (табл. 7).
Измерение силы условного рефлекса количеством слюны обнаруживает, что торможение в пункте задней лапы усилило условное возбуждение в пункте передней лапы почти на 50 %. Следовательно, в данном случае положительная индукция произошла из очага торможения в дальнем пункте анализатора.
Таблица 7. Положительная индукция из очага дифференцировочного торможения в кожном анализаторе (по Д.С. Фурсикову, 1922)
Однако мозг часто дифференцирует раздражители, связанные с одним и тем же пунктом анализатора, но отличающиеся друг от друга по силе или характеру воздействия. Будет ли в таких случаях проявляться положительная индукция? Ответ на этот вопрос дает следующий опыт. У собаки был выработан условный пищевой рефлекс на сильный свет, от него отдифференцировали слабый свет. Затем сильный свет был испробован сразу после слабого. И здесь условный слюнной рефлекс, вызванный сразу после дифференцировочного, увеличился почти на 50 %. Следовательно, в данном случае положительная индукция произошла из очага торможения в том же пункте анализатора.
Таким образом, положительная индукция может проявляться в различных анализаторах и при разных отношениях тормозного очага и положительно индуцируемого рефлекса.
Отрицательная индукция из очага возбуждения в очаг торможения. Явления отрицательной индукции можно продемонстрировать в следующем опыте. У собаки образован условный пищевой рефлекс на метроном 120 ударов/мин. К этому положительному раздражителю выработана дифференцировка метронома 60 ударов/мин. Как известно, дифференцировку очень легко разрушить, если начать сопровождать дифференцировочный раздражитель подкреплением. И действительно, после того как несколько раз метроном 60 ударов/мин применили с подкармливанием, он сам начал вызывать слюноотделение. Это простой и безотказный способ уничтожения тормозного очага.
Однако при помощи некоторых средств можно задержать разрушение дифференцировки, т.е. продлить существование очагов условного торможения. В частности, таким средством оказалось применение положительных сигналов, т.е. создание очагов условного возбуждения. Это видно из следующих опытов.
Например, собаку подкармливают после каждого применения метронома с частотой 60 ударов/мин до тех пор, пока у нее не начнет сильно выделяться слюна (разрушение дифференцировки). Тогда применяют с подкреплением один раз метроном с частотой 120 ударов/мин. В результате используемый вслед за ним метроном <...часть числа не видна…>0 ударов/мин, который только что вызывал слюноотделение, сразу теряет свое действие. Дифференцировка при этом восстанавливается, что связано с возникновением рядом очага возбуждения. Этот очаг отрицательно индуцировал, т.е. затормозил клетки пункта метронома с частотой 60 ударов/мин, и индукционное торможение усилило остатки дифференцировочного.