По мере дальнейшего развития дифференцировочного торможения неподкрепляемый сигнал при каждом своем применении будет усиливать колебания основного ритма и подавлять более быстрые колебания. При этом колебания основного ритма сами сильно замедлятся. Когда дифференцировка будет прочно выработана, отрицательный раздражитель — метроном с частотой 120 ударов/мин — не вызовет никакой двигательной реакции, но каждое его применение станет приводить к изменению электрической активности коры в сторону ее замедления. Часто при этом развивается дремотное состояние и возникают гигантские волны дельта-ритма. Однако медленные волны, наблюдаемые при угашении и выработке дифференцировок, рассматривают как необязательно связанные с развитием сна, а отражающие состояние неподвижности при бодрствовании, обозначая их «ритмами бодрствования — неподвижности». Отмечают, что при глубоком угашении условного рефлекса и выработке прочных дифференцировок медленные волны могут уменьшаться и электрическая активность возвращается к исходному состоянию.
Возможно, переделка ритмических свойств, т.е. подвижности корковых нейронов, составляет наиболее трудную работу торможения. Что касается гигантских размеров медленных волн, то они могут указывать на вовлечение в общий ритм «холостого хода» громадного числа нейронов, освобожденных торможением от разночастотных рабочих возбуждений. Медленные волны возникали при выработке запаздывающего условного рефлекса, при дифференцировании сигналов и во всех других случаях развития внутреннего торможения, что свидетельствует об их общей природе. Однако существуют указания, что в некоторых случаях тормозной раздражитель может не вызвать медленные волны, особенно в начале его действия.
Так как условные связи возникают в результате согласования пространственно-временной организации ритмической активности структур мозга (М.Н. Ливанов, 1970), то при торможении условного рефлекса происходит их рассогласование. Сходство электрических реакций коры на действие положительных и тормозных условных раздражителей привело к заключению, что на уровне коры и тот и другой вызывают процессы возбуждения, а торможение по их командам реализуется на уровне подкорковых исполнительных механизмов.
Таким образом, наиболее общее изменение в фоновой ритмике потенциалов при условном торможении — это сдвиг в сторону более редких ритмов, устранение бета- и резкое замедление альфа-ритма со значительным возрастанием амплитуды последнего. Тем самым электрические показатели характеризуют условное торможение как перестройку ритмических свойств корковых нейронов в сторону снижения их подвижности (лабильности).
Описаны изменения, которые претерпевали при развитии внутреннего торможения вызванные потенциалы. Однако в описании этих изменений существуют разногласия. Многие исследователи описывали уменьшение их величины, о которой судили главным образом по первичным ответам. Это отмечали при угашении условных рефлексов, например, на звуковые и зрительные раздражители при дифференцировании слуховых сигналов и при других видах внутреннего торможения. Другие авторы отмечали увеличение амплитуды вызванных потенциалов при угашении условных рефлексов у кошек и крыс, хотя в последнем случае дальнейшее углубление торможения приводило к снижению их амплитуды.
Вместе с тем накапливались факты, свидетельствующие о том, что первичные ответы, отражая общее состояние и уровень возбудимости коры, мало меняются при условно-рефлекторной деятельности. Поэтому особое внимание привлекли поздние компоненты вызванных потенциалов, которые считают наиболее близко отражающими процессы высшей нервной деятельности. Большинство авторов описывают их увеличение.
Из рис. 32 видны наступающее при углублении дифференцировочного торможения возрастание амплитуды первичного ответа и радикальная перестройка поздних компонентов вызванного потенциала. Многие авторы рассматривают усиление поздней поверхностно не<...часть слова пропущена...>тивной волны как дипольное отражение развития тормозных процессов в глубоких слоях коры. Уменьшение этих волн при подкреплении считают результатом блокирования тормозными клетками импульсации от ретикулярной формации среднего мозга. Однако известно и уменьшение поздних волн вызванных потенциалов. При развитии условного торможения изменяется не только амплитуда, но и форма поздних компонентов вызванных потенциалов, происходит перестройка положительных и отрицательных фаз и появятся новые компоненты.
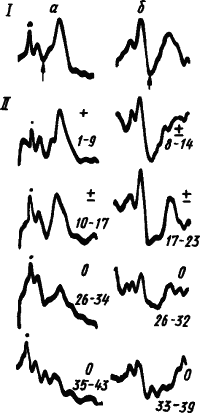
Рис. 32. Динамика вызванных потенциалов корковой проекции мышцы смыкания век на щелчки, подкрепляемые обдуванием глаза, при угашении условного рефлекса (по У.Г. Гасанову):
а, б — записи от двух подопытных животных; I — до угашения, II — в процессе угашения; арабскими цифрами указан порядковый номер подачи сигнала без подкрепления, знаки, означающие это сохранение условного рефлекса: «+» — частичное выпадение рефлексов, 0 — полное торможение, точки — артефакты от щелчков, стрелками отмечен пик поздней позитивной волны
К сожалению, пока не представляется возможным обобщить накопленные весьма разнородные сведения и дать им единое толкование. Видимо, неоднозначность результатов, особенно изменений поздних компонентов вызванных потенциалов, зависит от разных условий эксперимента: влияние посторонних раздражителей, локализации отведения, стадий развития условного торможения, фона электрокортикограммы, двигательной активности животного и многих других условий. В сложной картине динамики поздних негативных и позитивных компонентов вызванных потенциалов отражается развитие активности и смена состояния популяций нейронов разных слоев коры. Не менее сложные отношения складываются при регистрации вызванных потенциалов подкорковых структур. Вместе с тем систематический анализ динамики вызванных потенциалов, особенно его поздних волн, в разных структурах мозга при различных видах внутреннего торможения может быть ценным источником сведений о нейрофизиологическом механизме торможения.
Таким образом, при условном торможении изменения первичного ответа вызванного потенциала отражают главным образом общее состояние степени активности высших отделов мозга, а его поздние компоненты обнаруживают более специфические для развития тормозных процессов перестройки своей структуры, оценка значения которых требует дальнейших исследований.
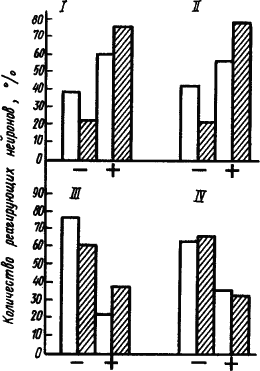
Так же как образование временной связи условного рефлекса происходило путем совместной деятельности возбуждающихся и тормозящихся нейронов, так и при развитии тормозного состояния в коре действуют как тормозящиеся, так и возбужденные нервные клетки (рис. 33). Видно, что при выработке дифференцировочного торможения в зрительной и двигательной зонах коры из общего числа реагирующих нейронов возбуждающихся оказывается даже больше, чем тормозящихся, а в лобных и теменных отделах заторможенных больше, чем возбужденных. Примечательно, что подобные реакции нейронов этих корковых областей отмечались и при образовании условного рефлекса. Так, максимальная тормозная реакция нейрона и ее латентный период совпадают с таковыми максимальной возбудительной его реакции при выработке условного рефлекса. Такое совпадение лишний раз подтверждает, что условное торможение развивается в тех же структурах, которые образовали временную связь, и морфофункциональные различия этих структур одинаково отражаются в свойствах как формирования временной связи, так и ее затормаживания. Преобладание возбужденных нейронов в зонах корковых проекций условного и безусловного раздражителей может указывать на высокую активность нервных процессов в этих «узловых» этапах условно-рефлекторной деятельности.