Параллельно цветковым растениям, млекопитающим и птицам меловую экосреду завоевывали костистые рыбы. Являясь отпрысками лучеперых рыб, предки которых зародились в период ордовикского оледенения, костистые пловцы были сильны в прохладной, малопродуктивной среде, поскольку лучше других рыбообразных создавали осевым скелетом двигательную локомоторную волну [124]. Недаром их современный рекордсмен, меч-рыба, развивает огромную скорость до 130 км/ч, превосходя по этой части наземного гепарда. Непреодолимо прогрессируя с раннемелового похолодания, они возобладали в морях с верхнего мела, легко пересекли мелтретичную границу и ныне составляют 95% ихтиофауны [86, с. 334; 655, с. 61–71]. Эта победа облегчалась малой мобильностью прочих мезозойских рыб [35, с. 205]. Эволюционная история акул, также имеющих ордовикские корни и отличающихся высокими ходовыми качествами, развивалась по образцу костистых рыб [86, с. 196–237] и тоже отмечена рекордами в кайнозое (см. ниже). Наконец, безуспешная борьба за жизнь медлительных аммонитов и белемнитов закончилась вытеснением их подвижными внутреннераковинными головоногими моллюсками: десятиногими кальмарами и каракатицами, а также и восьминогими осьминогами [35, с. 187; 84, с. 145–178].
С падением общеглобальной биопродуктивности среды разнообразие жизни на планете убывает, и проявления эволюционной направленности теряют разнообразие. Масштабы организмов тоже мельчают. Поэтому господствующие наземные животные мезозоя были крупнее кайнозойских, а раннекайнозойские – внушительнее позднекайнозойских современников нарастающего похолодания. Соответственно, примеры нашей версии эволюции для кайнозоя относительно скромны.
Кайнозой (65-0 млн лет назад)
В палеоцене (65–55 млн лет назад) высокомобильные организмы мелового периода продолжают разнообразиться, не меняя своей эволюционной направленности, поскольку вся эпоха отмечена регрессией и похолоданием. В начале эоцена (55–36 млн лет назад) началось метановое потепление [742], и регрессия сменилась трансгрессией. Следовало бы ожидать, что в установившихся теплых условиях появятся маломобильные формы мезозойского типа.
В соответствии с этим ожиданием в среднем эоцене распространяются уцелевшие потомки вымерших меловых белемнитов – байанотеутиды [35, с. 189–190; 38, с. 382, 388, рис. 217; 84, с. 161–162]. Тогда же возникают не связанные общностью происхождения нелетающие, бегающие птицы Ratidae, своего рода конвергенты (т. е. внешние подобия) двуногих динозавров: страусоподобные элеутерорнисы (средний эоцен, век лютет) и эпиорнисовые псамморнисы (эоцен) [35, с. 245–246]. Наконец, в раннем среднем эоцене Германии (50–48 млн лет назад) обнаруживается двуногое бегающее млекопитающее Leptictidium nasutum, подлинный конвергент двуногих динозавров со свойственным им и людям прямохождением (см. рис. 7) [118, с. 34]. Эоценовое потепление повысило биопродуктивность среды и раздвинуло рамки экониш, привлекая в них новых обитателей. В соответствии с этой новацией сухопутные парнокопытные произвели водных китообразных в нижне-среднем эоцене (ок. 52 млн лет назад [752]) или в конце среднего эоцена (в лютете, 47 млн лет назад [354]). В гармонии со своим происхождением в теплую эпоху китообразные (зубатые киты) развили чуждую млекопитающим равнозубость динозаврового типа.
В олигоцене (36–24 млн лет назад) трансгрессия прекращается и сменяется регрессией во второй половине эпохи. В связи с похолоданием высокомобильные четвероногие млекопитающие испытывают эволюционный подъем и порождают мегафауну, украшением которой является гигантский безрогий носорог Indricotherium transouralicum – самое крупное и высокое наземное млекопитающее, до 5 м высотой (средний олигоцен).
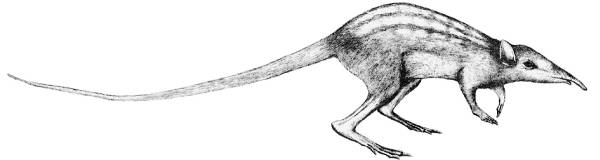
Рис. 7. Легкобег носатый (Leptictidium nasutum) возник в эпоху раннего среднего эоцена на островах нынешней Германии 50–48 млн лет назад, когда из-за потепления поднялась биопродуктивность среды, и естественный отбор дал шанс появлению маломобильных двуногих животных. «Носач» бегал на двух ногах, как двуногие динозавры и люди, а потому являлся их эволюционным конвергентом (подобием), таким же «теплолюбивым», как и они [118, с. 34]
В миоцене (24–5,3 млн лет назад) ранне-среднемиоценовое потепление (24–13 млн лет назад) положило конец олигоценовой мегафауне. Зато пробившийся отбор на маломобильность вызвал к жизни двуногую обезьяну «перелоапитека каталонского» (Испания, 13 млн лет назад), положившего начало целой плеяде двуногих гоминин (см. разд. 2. 4). Происходя от аборигенов высокобиопродуктивных тропических лесов, они органично несли в себе мезозойскую ориентацию на маломобильность и прожорливость, что в сочетании с эволюционной отсталостью создало гоминин, перспективных господ будущего, ставших лидерами именно в силу своей отсталости.
В плиоцене (5,3–1,6 млн лет назад) начало эпохи ознаменовалось трансгрессией. Затем последовал ряд гляциалов (оледенений), интергляциалов (межледниковий), стадиалов (пиков оледенений) и интерстадиалов (смягчений оледенений) выраженной ледниковой эпохи. Ее «лицом» можно назвать тридцатиметровую белую акулу Carcharodon megalodon, вымерщую в среднем плиоцене (3,3 млн лет назад), когда интергляциальная трансгрессия Астий вызвала ограниченное вымирание в среде мобильной биоты; cp. [680].
Наконец, в плейстоцене (1,6–0,0117 млн лет назад) развилась богатая приледниковая мамонтовая фауна, вымершая с началом голоценового потепления 11700 календарных лет назад, или 10200 лет назад по радиоуглероду 14С (поясним, что вследствие колебаний содержания тяжелого изотопа углерода 14С в древней атмосфере радиоуглеродные даты исследуемых образцов оказываются моложе своего реального, календарного возраста, который можно установить, например, методом дендрохронологии, когда высчитываются годичные кольца очень старых деревьев, до 8000 лет возраста, после чего итог сравнивается с радиоуглеродной оценкой; получается, например, что календарной дате 4650 лет назад отвечает радиоуглеродная дата 4050 лет назад).
Подобно динозаврам, мамонты и их спутники вымерли не сразу. Некоторые виды жили дольше: островные карликовые мамонты острова Сен-Пол в Беринговом море, 7908 ± 100 лет назад [391], островов Санта-Барбара в Калифорнии [480, с. 213], острова Врангеля на границе Восточно-Сибирского и Чукотского морей, 7620–3730 лет назад [515; 768]; большерогий олень в Западной Сибири, 6900 14C, или 7700 календарных лет назад [736]; возможно, шерстистый носорог в Чувашии, 921/922 гг. н. э. [54, с. 61–66, 139–140; 91, с. 72–74].
«Лицом» нынешней голоценовой эпохи – межледниковой, однако все еще не свободной ото льдов – можно считать гигантского кальмара Architeuthis longimana 19 м длиной (возможны тридцатиметровые особи), продолжающего линию внутреннераковинных головоногих моллюсков, которые сменили аммонитов в конце мелового периода.
Относительно упомянутых вымираний существуют неокатастрофические взгляды, касающиеся в первую очередь пермо-триасовых и мел-третичных событий. Согласно одному из них, массовые вымирания вызваны вулканической деятельностью планеты, которая обескислороживает океаны, отравляя их. Эти несчастья происходят с периодичностью в ~ 32 млн лет [632]. Они ответственны за массовую гибель биосферы на границе перми и триаса [227; 637; 639], а также мела и палеоцена [227; 598]; cp. [361, с. 368].
Показано, однако, что обескислороживание океанов господствовало на Земле на протяжении 250 млн лет в позднем неопротерозое ок. 800–543 млн лет назад [471; 802], однако на тогдашней жизни (например на морской эдиакарской фауне) это заметно не сказалось. Показано также, что обескислороживание океанов в самом начале триаса явилось следствием предшествующего позднепермского вымирания (кризиса радиолярий, морских микроорганизмов) [633], а никак не его причиной. Наконец, показано, что обескислороживание океанов имело место и в конце нижнего мела (в альбе, ок. 112 млн лет назад) [481], что не помешало костистым рыбам, акулам и прочим наследникам мезозоя уверенно набирать эволюционные обороты. В свете указанных расхождений с реальностью вулканическая гипотеза не может быть принята.