Основой бинокулярного стереоскопического зрения является диспаратность – оценка различия проекций изображений на сетчатках обоих глаз. Известно, что 84% нейронов зрительной коры бинокулярны. Они реагируют при одновременной стимуляции двух сетчаток, при этом нейроны одной колонки имеют близкие значения диспаратности. Благодаря тому, что правый и левый глаза рассматривают один и тот же объект с разных точек, изображения объекта на правой и левой сетчатке сдвинуты относительно друг друга. Это различие изображений зависит от того, насколько приближен или отдален объект относительно точки пересечения оптических осей двух глаз (точка фиксации). Нейрофизиологической основой бинокулярного зрения является взаимодействие двух рецептивных полей, которые имеются у каждого бинокулярного нейрона зрительной коры (на контра- и ипсилатеральной сетчатке). Каждый бинокулярный нейрон избирателен к какой-то одной определенной диспаратности (это определяется диспаратностью его рецептивных полей), а поскольку диспаратность зависит от удаленности объекта, реакции бинокулярных нейронов оказываются избирательными к определенной удаленности. В коре имеется целый набор нейронов с разной диспаратностью. Эта совокупность нейронов составляет механизм, измеряющий удаленность объекта. Описанный выше механизм диспаратности лежит в основе стереоскопического зрения.
Цветооппонентные нейроны в зрительной коре обезьян. Значительная часть цветооппонентных нейронов зрительной коры приматов имеет простые или концентрические рецептивные поля. Большинство нейронов с цветооппонентными рецептивными полями встречаются в слое IV. Для нейронов этого класса характерна цветовая оппонентность в пределах центра рецептивного поля: нейрон реагирует возбуждением на стимуляцию одного цветоприемника в центре рецептивного поля и тормозится при стимуляции другого. Одни нейроны реагируют on-ответом на красное освещение и off-ответом – на зеленое, реакция других – обратная. Белый свет не вызывает реакции этих нейронов, так как антагонистические влияния от двух цветоприемников взаимно погашаются. У нейронов с концентрическими рецептивными полями, помимо оппонентных отношений между цветоприемниками, существуют также обычные для концентрических полей антагонистические отношения между центром и периферией. В результате возникает структура с двойной цветооппонентностью. Если воздействие на центр рецептивного поля вызывает, например, on-ответ на красное освещение и off-ответ – на зеленое, то нейрон с такими свойствами сочетает избирательность к цвету с избирательностью к локальным изменениям яркости пятна соответствующего цвета. Эти нейроны не реагируют ни на стимуляцию белым светом (из-за оппонентных отношений между цветоприемниками), ни на диффузную стимуляцию светом любой длиной волны (из-за антагонистических отношений между центром и периферией рецептивного поля). Максимум реакции регистрируется, если одновременно стимулировать разные цветоприемники в центре и на периферии рецептивного поля. Цветооппонентные нейроны, избирательные к ориентации, могут иметь простые и сложные рецептивные поля. В простом рецептивном поле различают две или три параллельно расположенные зоны, между которыми имеется двойная оппонентность: если центральная зона имеет on-ответ на красное освещение и off-ответ на зеленое, то краевые зоны дают off-ответ на красное и on-ответ на зеленое. В сложном рецептивном поле нет раздельных зон с разной спектральной чувствительностью. Нейроны этого типа реагируют только на определенным образом ориентированные линии предпочитаемого цвета или цветоконтрастные границы. Большинство цветооппонентных нейронов коры у обезьян связано с красно- и зеленочувствительными цветоприемниками. С синечувствительными цветоприемниками связано значительно меньше нейронов. Предполагается, что информация, выделенная нейронами первичного зрительного поля 17, далее передается для обработки во вторичную (поле 18) и третичную (поле 19) области коры.
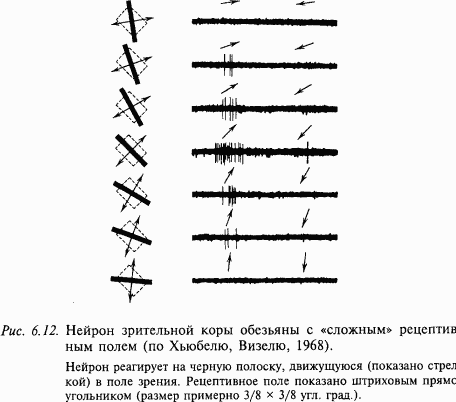
Колончатая организация зрительной коры. При погружении микроэлектрода перпендикулярно поверхности зрительной коры, как и в других корковых проекционных зонах (моторной, слуховой и т.д.), по ходу электрода всегда встречаются нейроны с близкими свойствами. Часто эти нейроны объединяются по признаку организации своих рецептивных полей: в одной колонке собраны нейроны примерно с одной ориентацией рецептивных полей. Упорядоченность в расположении ориентационных колонок очень высока в зрительной коре обезьян. Смещение регистрирующего микроэлектрода в коре на каждые 25–50 мкм в тангенциальном направлении приводит к повороту рецептивного поля регистрируемых нейронов в одном и том же направлении (по часовой стрелке или против) на величину примерно 10°, так что полный набор колонок со всеми ориентациями рецептивных полей в пределах 180° занимает в коре участок 500–1000 мкм. В пределах этого участка отдельные колонки в зрительной коре обезьяны в поперечном (параллельно поверхности коры) сечении имеют вид узких полосок шириной 25–50 мкм. Под гиперколонкой понимается участок коры, включающий набор ориентированных колонок (с ориентациями в пределах 180° и перекрывающиеся с ними две окулодоминантные колонки). Такая гиперколонка обрабатывает информацию от определенного участка сетчатки; информация от соседних участков обрабатывается такими же соседними гиперколонками. Однако из вышеизложенного не следует, что нейроны, составляющие колонку, в функциональном отношении совершенно однотипны. Например, в зрительной коре кошки одна колонка содержит нейроны с простыми и сложными рецептивными полями. В этом случае общим признаком будет только ориентация рецептивных полей отдельных нейронов. В одной колонке могут находиться нейроны с одинаковой цветоизбирательностью и другими однотипными свойствами. Обнаружено, что упорядоченность в размещении нейронов зрительной коры обезьян имеет место не только по вертикали в виде колонок, но и по горизонтали. Так, например, нейроны с простыми рецептивными полями находятся преимущественно в глубине III и IV слоев. Нейроны со сложными и сверхсложными рецептивными полями локализованы в основном в верхних (II и верхняя часть III) и нижних (V и VI) слоях коры.
Способен ли новорожденный видеть зрительный мир так, как его видит взрослый? Эксперименты на животных показали, что многие свойства зрительной системы заложены с рождения, но развитие этих способностей зависит в дальнейшем от воспитания. Например, было установлено, что у 1–2-недельных котят избирательные свойства корковых нейронов уже сформированы, т.е. они образовались исключительно за счет генетических программ в отсутствие зрительных стимулов (котята в этот период еще слепые). Вместе с тем если у котенка в течение первых 2–3 мес. жизни исключить предметное зрение, например, одним глазом, то нейроны коры, соответствующие этому глазу, теряют ориентированные зрительные поля. Интересно, что свойства нейронов наружного коленчатого тела при этом заметно не страдают. Отсюда был сделан вывод, что свойства корковых нейронов определяются исключительно внутрикорковыми связями. У котят период, чувствительный к депривации, начинается после 4–5 недель после рождения и длится до 6–8 недель. У низших обезьян этот период значительно дольше: начинается через 1–2 мес. после рождения и продолжается до 1,5–2 лет. Интересные результаты были получены в экспериментах с воспитанием котят в определенной зрительной среде. Например, если котята в течение первых месяцев после рождения видят только вертикальные (черно-белые) или только горизонтальные полосы, то в более поздние сроки в зрительной коре обнаруживаются нейроны с ориентацией рецептивных полей, соответствующих зрительной среде периода воспитания. Из этих экспериментов было сделано два важных вывода: 1) межнейронные связи, соответствующие данному виду животного, программируются генетически; 2) в то же время в раннем онтогенезе имеется период, чувствительный к воздействию внешних условий среды обитания данного вида. Такое двойное обеспечение межнейронных связей представляется биологически целесообразным.