Большое число рецепторов спонтанно разряжаются в отсутствие каких-либо раздражителей. Например, рецепторы вестибулярного аппарата постоянно активны, что дает им возможность сигнализировать не только о величине, но и о направлении действующего стимула (например, ускорения). Для этих случаев порог определяется не как возникновение, а как изменение реакции рецептора – дифференциальный порог.
При длительном действии адекватного стимула порог срабатывания данного рецептора повышается. Это явление называется адаптацией. В адаптации рецептора принимает участие большое число процессов, и ее нельзя объяснить каким-либо одним механизмом. Например, в рецепторах кожи самым важным местом адаптации является ткань, окружающая нервное сенсорное окончание. К этой механической адаптации прибавляется адаптационный процесс генераторного потенциала. В фоторецепторах сетчатки также происходит несколько процессов: одни связаны с фотохимическими реакциями, другие – с активностью нервной сети сетчатки. В каждом рецепторе имеется свое сочетание факторов, определяющих процесс адаптации.
Отдельное нервное афферентное волокно 1-го порядка собирает информацию с более или менее широкой области поля рецепторов. Та часть рецепторной поверхности, от которой сигналы получает одно афферентное волокно, называется его рецептивным полем. Рецептивные поля соседних элементов, как правило, перекрываются. Перекрытие отдельных рецептивных полей, по-видимому, играет значительную роль. Прежде всего, этим обусловливается надежность функции, которая благодаря такой организации обеспечивается большим числом рецепторов.
Одним из важных принципов взаимодействия в нейронных сетях является латеральное торможение: возбужденные элементы (рецепторы, нейроны) через возвратные коллатерали затормаживают соседние элементы. Известно, что латеральное торможение существует на всех уровнях сенсорных систем. Благодаря такому тормозному взаимодействию предотвращается «растекание» возбуждения по нервной сети, происходит своеобразное увеличение контраста, т.е. степени перепада между возбужденными и невозбужденными нейронами.
В состав сенсорной системы, кроме описанного 1-го уровня обработки информации, входит также ряд других структур головного мозга с соответствующими проводящими путями. Одними из важных станций переключения афферентной импульсации к коре служат специфические ядра таламуса. Высшим уровнем обработки сенсорных сигналов является кора больших полушарий, которая достигает наибольшего развития у млекопитающих и особенно у приматов. Кора головного мозга млекопитающих, покрывающая большие полушария конечного мозга, благодаря многочисленным складкам может иметь значительную площадь; у человека, например, она достигает 1700–2500 см2. В начале XX века была разработана классификация полей коры по признакам особенностей клеточного состава и характера миелинизации аксонов. На основании изучения клеточного состава отдельных слоев в коре выделено 11 областей, которые, в свою очередь, были разделены на 52 поля (рис. 6.4).
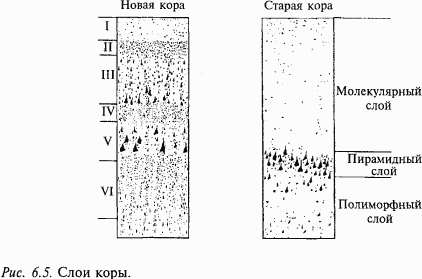
Толщина коры у млекопитающих разных видов колеблется от 1 до 6 мм. Нейроны по толщине коры распределены неравномерно и обычно образуют в новой коре 6 слоев, в старой коре – 3 слоя (рис. 6.5). Однако в отдельных областях новой коры количество слоев может увеличиваться или уменьшаться. Слои V и VI содержат преимущественно крупные пирамидные нейроны, аксоны которых образуют эфферентные пути из коры. Основные афферентные пути в кору заканчиваются на нейронах верхних слоев (III и IV). Эти слои наиболее сильно развиты в центральных отделах зрительного, слухового и кожного анализаторов. По современным представлениям, такое деление слоев коры на афферентные и эфферентные нужно считать в значительной степени условным. В последние годы при изучении внутрикорковых связей установлено, что эфферентные аксоны нейронов нижних слоев образуют многочисленные возвратные коллатерали, которые восходят до самых верхних слоев коры.
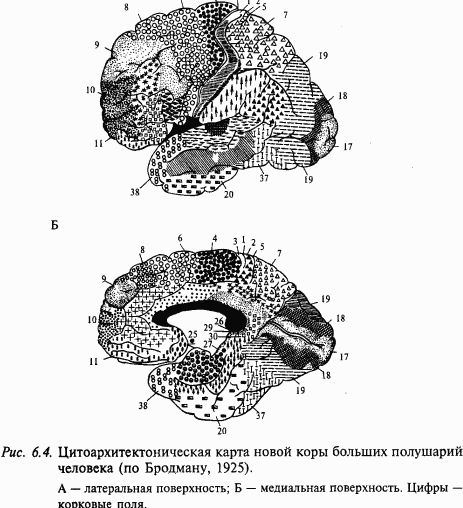
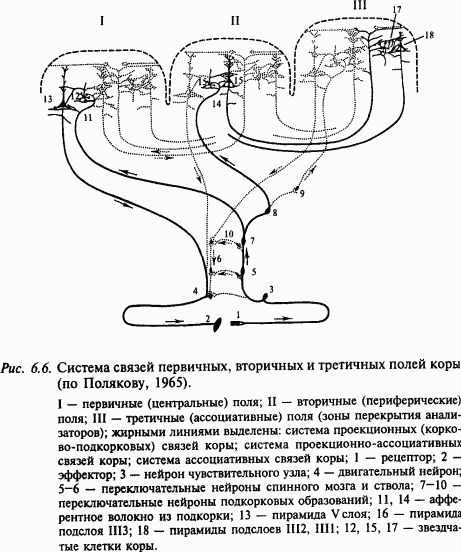
В соответствии с цитоархитектоническими и нейрофизиологическими данными выделяют проекционные и ассоциативные зоны коры. В проекционных зонах различают первичные, вторичные и третичные зоны. По И. П. Павлову, под ядром анализатора понимают зону коры, в которой после соответствующих переключений в подкорковых ядрах имеет место детальное представительство периферического отдела анализатора. Принцип деления на первичные, вторичные и третичные поля иллюстрируется рис. 6.6, на котором схематично показано, что возбуждение от соответствующих рецепторов направляется в первичные зоны по быстропроводящим путям, тогда как активация вторичных и ассоциативных зон коры происходит по полисинаптическим путям. Кроме этого, корковые поля связаны между собой многочисленными ассоциативными волокнами. В корковых проекциях сенсорных систем соблюдается принцип топии, например, в зрительной системе ретинотопия – проекция сетчатки – в поле 17 (первичная проекция), поле 18 (вторичная) и поле 19 (третичная). Этот принцип состоит в том, что, например, сетчатка, образует проекцию на соответствующую область коры в некотором масштабе. При этом объем корковой проекции пропорционален плотности рецепторов в данном месте сетчатки. Благодаря этому fovea (зрительная ямка) сетчатки в корковой проекции представлена большей площадью, чем периферия сетчатки. Поражение первичных зон сенсорных систем всегда сопровождается грубыми нарушениями соответствующей сенсорной или моторной функции. В отличие от этого поражение вторичных или третичных зон вызывает расстройство аналитико-синтетической деятельности мозга, например гнозиса и праксиса, речи, целенаправленного поведения и пр. Подробнее см. главу 7 «Высшие функции нервной системы».
Вопросы
1. Принцип разделения рецепторов на первичные и вторичные.
2. Типы сенсорных порогов.
3. Виды кодирования сенсорного стимула в сенсорной системе.
4. Строение сенсорной системы.
Литература
Батуев А. С., Куликов Г. А. Введение в физиологию сенсорных систем. М.: Высшая школа, 1991.
Основы сенсорной физиологии./Под ред. Р. Шмидта. М.: Мир, 1984.
Сомьен Дж. Кодирование сенсорной информации. М.: Мир, 1976.
Шульговский В. В., Ерченков В. Г. Сравнительная физиология анализаторов. М.: Изд-во Моск. ун-та, 1989.
НЕЙРОФИЗИОЛОГИЯ ЗРИТЕЛЬНОЙ СИСТЕМЫ
Зрение в жизни человека имеет огромное значение. Это основной сенсорный канал, который связывает его с внешним миром. Зрительная система человека устроена очень сложно. Благодаря зрению мы воспринимаем окружающий нас мир в объеме и красках, мы читаем и смотрим кино, телевизор. И это все зрение.
В зрительной системе человека можно выделить следующие уровни обработки сигналов. На периферии находится сетчатка. В ходе развития нервной системы сетчатка закладывается на самых ранних этапах развития (так называемые «глазные пузыри»). Поэтому есть все основания считать сетчатку «частью мозга, вынесенного на периферию». Следующий уровень обработки зрительной информации находится в таламусе – это наружное коленчатое тело. Аксоны нейронов наружного коленчатого тела проецируются в кору затылочного полюса больших полушарий (поля 17, 18, 19). Высший этап обработки зрительных сигналов происходит в ассоциативных полях коры больших полушарий. Схема зрительной системы приведена на рис. 6.7.
Строение глаза. Глаз человека имеет шарообразную форму (рис. 6.8). Вращение глазного яблока в глазнице осуществляется тремя парами мышц, которые иннервируются глазодвигательными черепными нервами (подробнее см. выше). Плотная наружная оболочка глаза образована непрозрачной склерой, которая на переднем полюсе переходит в прозрачную роговицу. Внутри глазного бокала находится сосудистая оболочка, содержащая кровеносные сосуды. Впереди сосудистая оболочка переходит в ресничное тело и далее в радужку. В радужке находятся гладкие мышечные волокна, степень напряжения которых определяет диаметр зрачка. При сокращении или расслаблении гладкой мускулатуры ресничного тела изменяется напряжение цинновых связок, от которых зависят радиус кривизны хрусталика и его преломляющая сила, т.е. аккомодация глаза. Пространство между хрусталиком и роговицей, называемое передней камерой, заполнено прозрачной жидкостью, между хрусталиком и сетчаткой – студенистой жидкостью, или стекловидным телом. Дно глазного бокала выстлано сетчаткой.