Таким образом, у данных видов стрекоз существует вполне развитое территориальное поведение: занимаются индивидуальные участки, в которых выделяются функциональные зоны (отдыха, размножения), на этих участках выполняются все жизненные функции (за исключением сна: вечером самцы собираются в особых местах ночевок), хозяин дает знать о своем присутствии, маркирует и активно защищает участок.
Интересные особенности территориального поведения муравьев были выявлены советским энтомологом А. А. Захаровым. Оказалось, что у муравьев существуют два основных типа использования кормовых участков: совместное использование угодий несколькими семьями и использование кормового участка населением лишь одного гнезда. При этом выявилась прямая связь между плотностью муравьев на участке и «агрессивностью»: у видов с низкой плотностью участки не охраняются (кроме пригнездовой зоны), при высокой же плотности на кормовых участках появляются охраняемые территории, а между ними «нейтральные зоны» (рис. 40). Как показал другой советский исследователь, Г. М. Длусский, муравьи того же вида, принадлежащие к другим семьям, равно как и представители других видов, в пределы этих территорий не допускаются.
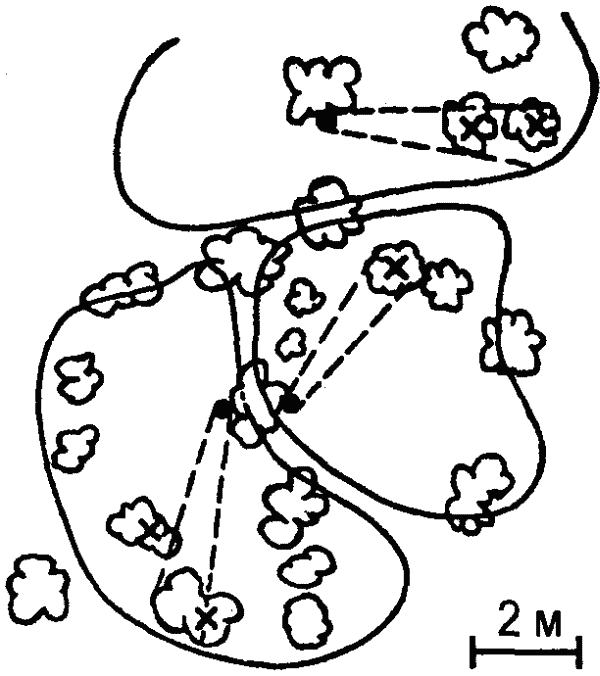
Наибольшую сложность территориальное поведение достигло у рыжих лесных муравьев, у которых наблюдается и наибольшая плотность особей на кормовых участках. Охраняемые территории этих муравьев достигают значительно большей величины, чем у других видов. Центральными элементами такой территории являются муравьиные тропы (постоянные кормовые дороги), длина которых нередко превышает 100 метров, а протяженность всей сети троп — 1 километр. Каждой дорогой пользуется лишь определенная группа муравьев («колонна»), занимающая в муравейнике определенный сектор, примыкающий к «своей» дороге. Территория всей семьи подразделяется дорогами на отдельные части, составляющие территории отдельных колонн. Между ними имеются такие же нейтральные пространства, как и между территориями семей, хотя и не столь выраженные, как в последнем случае. Границы территории маркированы и препятствуют «рассеиванию» муравьев. Муравьи двигаются по территории не хаотично, а в соответствии со структурой территории: в центральной и средней ее частях — по кратчайшему пути от дорог или муравейника, на периферии же — параллельно границам территории. Таким образом, траектория движения каждого муравья детерминируется местом нахождения его на территории (исследования И. В. Стебаева). Наибольшую охотничью активность муравьи развивают на периферии территории (далее 10 метров от гнезда). Сбор строительного материала производится в пределах двухметровой полосы вдоль дорог.
Захаров справедливо отмечает сходство территориального поведения муравьев с таковым у птиц и хищных млекопитающих.
Инстинкт и научение в поведении насекомых
Долгие годы господствовало мнение будто насекомые и другие членистоногие являются существами, поведением которых руководит жесткий «слепой инстинкт». Это представление укоренилось преимущественно под влиянием работ выдающегося французского энтомолога Ж. А. Фабра, который сумел своими блестящими исследованиями убедительно показать, что даже сложнейшие действия насекомых не являются проявлением «разума», а выполняются на врожденной, инстинктивной основе. Одностороннее развитие положений Фабра и привело к указанной, неверной оценке поведения насекомых, к отрицанию не только разумности их поведения, но и к отрицанию, или хотя бы умалению роли накопления индивидуального опыта, научения в их жизни.
Как мы уже видели, формирование любой формы видотипичного, наследственно «закодированного», т. е. инстинктивного, поведения в онтогенезе всегда сопряжено в той или иной степени с какими-либо элементами индивидуально приобретаемого поведения, научения. О строго фиксированном инстинктивном поведении в «чистом виде» не приходится говорить даже относительно низших животных.
В полной мере это относится и к насекомым, инстинктивное поведение которых также совершенствуется научением. В этом состоит основная роль научения в жизни насекомых. Можно, очевидно, считать, что научение стоит у насекомых и других членистоногих «на службе» у инстинктивного поведения. Как и у других животных, инстинктивные движения (врожденные двигательные координации) у них генетически строго фиксированы. Инстинктивные же действия, инстинктивное поведение являются и у насекомых в той или иной степени пластичными благодаря включению в них благоприобретаемых компонентов.
В естественных условиях способность к накоплению индивидуального опыта проявляется у насекомых в неодинаковой степени в разных функциональных сферах. Чаще всего она связана с ориентацией в пространстве и пищедобывательной деятельностью. Примером могут служить отмеченные выше опыты по обучению пчел за пищевое подкрепление ориентироваться по различным рисункам. Другой пример — муравьи, которые очень легко (всего за 12–15 опытов) научаются проходить даже сложный лабиринт, но, насколько известно, не научаются действиям, лежащим за пределами указанных функциональных сфер. Такая специфическая направленность (и одновременно ограниченность) способности к научению является характерной особенностью научения у представителей всего типа членистоногих.
Роль научения в поведении насекомых наглядно выступает и в «танцах» пчел — этих высших представителей членистоногих. Отстаивая взгляд, будто насекомые, в том числе пчелы, являются «стимульно связанными, рефлекторными животными», американские ученые В. Детьер и Э. Стеллар заявляют, например, что выполнению и интерпретации сложного танца пчелы не обучаются. Вместе с тем, как показали советские исследователи Н. Г. Лопатина, И. А. Никитина, Е. Г. Чеснокова и другие, процессы научения не только уточняют, но и модифицируют коммуникационные способности пчелы в онтогенезе и расширяют набор сигнальных средств.
Более того, как установили названные исследователи, биологическая значимость сигнальной деятельности медоносных пчел определяется стереотипом условных рефлексов, приобретаемых в онтогенезе по мере освоения пространства и при общении в семье. Оказалось, что интерпретация передаваемой в танце информации о расстоянии и направлении полета к источнику пищи возможна лишь в том случае, если пчела до этого научилась соотносить местонахождение корма с характером информации, содержащейся в танце сборщиц. Кроме того, тактильный компонент танца (вибрации брюшка) не имеет врожденного сигнального значения. Последнее приобретается в онтогенезе также условно-рефлекторным путем: пчелы, не имевшие в онтогенезе контактов (пищевых) с танцовщицей, не в состоянии интерпретировать этот существеннейший элемент танца. Следовательно, каждая пчела должна в основном научиться «понимать» язык танца. С другой стороны, образование временных связей оказалось важным и для формирования самой способности к выполнению танцев.
Таким образом, нет неизменных форм поведения даже там, где прежде всего требуется стереотипность, — в сигнальных позах и телодвижениях. Даже такое врожденное коммуникативное поведение, как «танцы» пчел, не только дополняется и обогащается процессами научения, не только переплетается с ними, но и формируется в комплексе с индивидуально приобретаемыми элементами поведения.