Так вот, возвращаясь к появлению в триасе разнообразных морских рептилий (кстати: именно тогда сделали попытку освоить море и амфибии-лабиринтодонты). Многие исследователи (например, Р. Кэрролл) полагают, что переход к водному образу жизни происходит у низших амниот особенно легко именно в силу низкой интенсивности их метаболизма, способности выдерживать недостаток кислорода (из-за использования анаэробных процессов для работы мышц) и низкой температуры тела (не выше, чем у окружающей воды). Иными словами – рептилии исходно предрасположены к обитанию в водной среде. Судя по наблюдениям над современными морскими игуанами, передвижение и питание в воде не требует от них структурной или физиологической перестройки; более того – при водной локомоции метаболические затраты этих ящериц составляют лишь четверть от тех, что потребны для перемещения по суше (благо тело в воде ничего не весит – по закону Архимеда)[58] . Рептилии без проблем становятся вторичноводными всякий раз, когда жить в воде становится выгоднее с точки зрения обилия пищи и/или отсутствия врагов и конкурентов. Так, в юре возникли морские крокодилы (ноги их превратились в настоящие ласты, и имелся специальный хвостовой плавник, как у ихтиозавра), в раннем мелу – морские черепахи, близкие к нынеживущим, а в позднем мелу – мозозавры, исполинские (до 12 м длиной) морские ящерицы-вараны (рисунок 42, д-е).
На суше ситуация складывется принципиально иначе, чем в море: тут белая мускулатура создает для завроморфов серьезные проблемы. Мы уже упоминали о различиях в строении поясов конечностей у тероморфов и завроморфов. Конечности могут быть расположены сбоку от тела (такое их положение называется латеральным), или подведены под него (парасагитально); в первом случае бедренная (или плечевая) кость направлена параллельно земле, а во втором – вертикально (рисунок 43, а). Отсюда следуют различия в типе локомоции: рептилии «враскорячку» ползают на брюхе (отсюда происходит русское название группы – «пресмыкающиеся»), тогда как у маммалий тело высоко приподнято над землей, а конечность, двигаясь в плоскости, параллельной оси тела, способна делать широкий шаг (рисунок 43, б). Весьма существенно то, что при парасагитальном положении конечностей (маммальный вариант) вес тела принимают на себя – продольно! – кости скелета конечностей, так что мышцам надо лишь не давать этим «подпоркам» отклоняться от вертикального положения; при латеральных же (рептилийных) конечностях тело «подвешено» на самих мышцах, и животное как бы постоянно выполняет упражнение «отжимание от пола» – удовольствие ниже среднего.
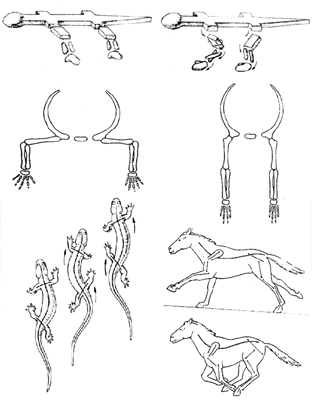
РИСУНОК 43. а – Положение конечностей в латеральном и парасагитальном положении; б – «рептильный» и «маммальный» шаг.
Казалось бы, маммальное строение поясов конечностей безусловно «лучше». В действительности же свои плюсы есть и в рептилийном варианте: ведь если мускулатура у тебя по преимуществу белая, ориентированная «на рывок», то проще большую часть времени проводить, расслаблено лежа на брюхе, и лишь изредка совершать броски за добычей. При маммальной же постановке конечностей приходится постоянно поддерживать тело на весу; это во-первых практически исключает саму возможность использовать белую мускулатуру (которая «устает»), а во-вторых, заставляет тратить много энергии вхолостую – мы помним, что млекопитающее в покое расходует в покое примерно в 10 раз больше энергии, чем рептилия равного размера. Однако когда все плюсы и минусы просуммированы, оказывается, что на суше для завроморфов (с их белой мускулатурой и латеральной конечностью) открыты лишь «профессии» подстерегающего хищника или пассивно защищенного – панцирем или слоем воды – крупного фитофага; остальные экологические ниши оказываются в распоряжении «теплых», активных, высокоподвижных тероморфов.
Ситуация поменялась в триасе, когда на эволюционную арену вышла группа рептилий, являющая собою подлинный венец всей завроморфной линии эволюции – архозавры. К этому подклассу принадлежат крокодилы и вымершие текодонты, динозавры и птерозавры; прямо от него ведут свое происхождение птицы. Из курса зоологии вы помните, что архозавры (о строении которых мы судим в основном по крокодилам) обладают целым рядом уникальных для рептилий продвинутых черт: у них уже полностью разделены правый и левый желудочки сердца, зубы их альвеолярного типа – сидящие в отдельных ячейках (что позволяет аллигаторам иметь «клыки» до 5 см длиной), возникает вторичное нёбо – дополнительная костная перемычка под первичным нёбом, отделяющая дыхательные пути от ротовой полости. Однако для триасовой победы архозавров над тероморфами наиболее существенным оказалось, по всей видимости, именно видоизменение конечностей и их поясов (в этом смысле крокодилы как раз не показательны – их конечности очень примитивны, близки к предковому для архозавров типу), которое весьма неожиданным образом разрешило проблему скоростной локомоции.
Дело в том, что по ряду анатомических причин (они были детально обсуждены еще А. Ромером в 1922 г.) перевести в парасагитальное положение задние конечности, которые причлененны к позвоночнику, намного проще, чем передние, соединенные с грудиной. Достаточно сказать, что тероморфы сумели «подвести под тело» задние ноги еще в перми (у горгонопсид), а вот полная парасагитальность выработалась в этой линии много позже – лишь у триасовых цинодонтов. Итоговая же скорость передвижения «до-цинодонтного» тероморфа, как легко догадаться, все равно лимитируется именно «медленными» – латеральными – передними конечностями.
Архозавры появивлись в поздней перми, почти одновременно с терапсидами, и одновременно же с последними выработали парасагитальность задних конечностей. А вот дальше архозавры, вместо того, чтобы возиться с преобразованием пояса передних конечностей, вообще отказались от их использования для движения, и выработали принципиально новый тип локомоции – бипедальный (двуногое хождение); о потенциальных скоростных возможностях такого двуногого диапсида дают некоторое представление современные страусы... Бипедальность тоже требует ряда серьезных анатомических перестроек (прежде всего – создания опорного таза с консолидированными позвонками крестцового отдела), однако это, как ни странно, оказалось легче, чем решить простенькую на первый взгляд задачу снятия ограничений в подвижности плечевого сустава – на что тероморфам пришлось потратить почти 100 миллионов лет[59] .