Другой неожиданный вывод следует из изучения открытых в конце восьмидесятых годов в Швеции среднекембрийских фосфатных конкреций Ёрстен. При тамошнем типе сохранности фиксируются не покровные ткани (как в Бёждес), а микроскопические щетинки и иные твердые покровные элементы, видимые лишь под сканирующим электронным микроскопом. Оказалось, что в то время существовал многочисленный и очень разнообразный микрозоопланктон – трохофоры (личинки кольчатых червей), зоеа (личинки ракообразных) и т.п. Это позволяет вновь вернуться к гипотезе А.Ю.Розанова (1986) о том, что в кембрии в действительности произошло лишь увеличение размеров уже существовавших на тот момент планктонных организмов, которые по этой причине «выпали в осадок» – перешли к бентическому образу жизни; дополнительные аргументы обретают и построения М.Б. Бурзина (см. выше) о эволюции планктонных экосистем.
Надо заметить, что мир первых скелетных организмов, сложившийся в начале кембрия (немакит-далдынский ярус), был крайне своеобразен и в чистом виде просуществовал очень недолго, лишь до середины этого периода – хотя отдельные ого представители дожили до середины ордовика. В это время уже существовали достаточно многочисленные животные, имевшие раковины (моллюски и брахиоподы) и жилые трубки (сабелидиты, хиолиты) и т.п.; разнообразнейшие мелкие зубчики – конодонты представляли собою части ротовых органов древнейших вторичноротых – щетинкочелюстных и хордовых. Весьма характерны для этого мира приапулиды (в современных морях эта реликтовая группе насчитывает лишь 20 видов), а также лобоподы (представленные ныне микроскопическими тихоходками и обитающими в подстилке влажных тропических лесов онихофорами); видимо, именно к лобоподам принадлежат многие загадочные кембрийские организмы, вроде галлюциногении (оцените название!). Имелись и и многочисленные артроподоподобные формы (см. ниже) (рисунок 25, а-д).
Основу тогдашних экосистем, однако, составляли водоросли, строившие небольшие известковые постройки – биогермы, и чрезвычайно любопытная, как считалось, вымершая группа организмов – археоциаты. Археоциаты (по-гречески – «древние кубки») представляют собой небольшие прикрепленные ко дну бокалы с двуслойной стенкой (рисунок 24); диаметр их колеблется от нескольких миллиметров до нескольких сантиметров, хотя в Сибири найдены и исполинские формы размером до 1,5 м. Природа их долго вызывала споры (неясно было даже – животные это или растения, или вообще отдельное царство), однако в последнее время почти все исследователи согласны с тем, что их следует относить к губкам. Более того – не так давно в Тихом океане была найдена странная глубоководная губка Vacletia, оказавшаяся по рассмотрении дожившим до наших дней археоциатом и пополнившая собою галерею «живых ископаемых» (вроде кистеперой рыбы или гинкго).
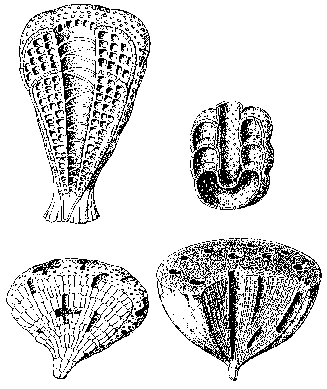
РИСУНОК 24. Археоциаты. (а) – Archaeocyatida, (б) – Capsulocyatida, (в) – Kazachstanicyatida, (г) – Archaeocyatida.
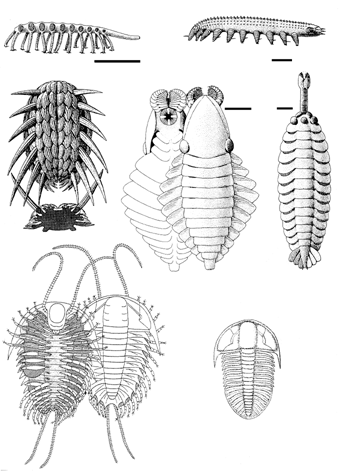
РИСУНОК 25. Различные кембрийские животные: лобоподы (а-б), члнистоногие неясного систематического положения (в-д), трилобиты (е-ж). (а) – Xenusion; (б) – Aysheaia; (в) – Wiwaxia; (г) – Anomalacaris; (д) – Opabinia; (е) – Olenoides , (ж) -Ogygopsis.
Губки занимают совершенно обособленное положение в системе животного царства и могут быть противопоставлены всем остальным многоклеточным животным: есть веские основания считать, что они ведут свое происхождение от иной группы простейших, нежели прочие метазоа. Существует, например, остроумная и неплохо аргументированная гипотеза Й. Райтнера (1991), согласно которой губки возникают как симбиоз бактериальной колонии с хоанофлагеллятными простейшими[22] . Достигнутый губками уровень организации не позволяет назвать их многоклеточными в строгом смысле («многотканевыми» по Корлиссу), так что вроде бы нет ничего удивительного в том, что именно эта примитивнейшая группа животных открывает собою фанерозойскую летопись. Но это только на первый взгляд.
Дело в том, что крайне низкий уровень клеточной интеграции оставляет губкам единственно возможную «профессию» – прикрепленный пассивный фильтратор органической взвеси. Любая губка (включая археоциат) представляет собою открытый сверху мешок, стенки которого пронизаны каналами; вода постоянно движется сквозь них во внутреннюю (парагастральную) полость, выходя затем наружу через верхнее отверстие (устье), а входящие в состав стенок каналов хоаноциты (воротничковые жгутиковые клетки) отфильтровывают содержащуюся в воде органику и бактерий. Вопрос для школьной олимпиады: какая сила заставляет воду двигаться сквозь каналы в неподвижной стенке? Ответ: та же самая, что создает печную тягу – разница давлений на высоте поддувала и на высоте конца трубы в соответствии с законом Бернулли[23] : если водная среда, в которой находится губка, имеет ненулевую скорость, то слой движущейся жидкости над устьем создает «подсос». Известное дело: чем выше труба – тем лучше тяга; соответственно, губка может прокачивать сквозь себя воду лишь когда ее устье приподнято над субстратом. (Сходный механизм обеспечивает вентиляцию нор сусликов и иных грызунов. Тот из выходов, через который при рытье норы выбрасывали грунт, оказывается заключен в холмик-"кротовину" и несколько приподнимается над землей; в итоге внутри норы возникает устойчивый ток воздуха от «низкого» выхода к «высокому»). Подъем же устья над субстратом можно обеспечить лишь при наличии твердого опорного скелета – органического или минерального (у современных губок есть оба варианта). Отсутствие в предшествующих отложениях этих легко обнаруживаемых скелетных элементов – спикул – свидельствует о том, что данная группа живых организмов действительно возникла лишь в начале кембрия (возможно, именно по схеме Райтнера – см. выше), т.е. когда уже несомненно существовали и настоящие многоклеточные животные.
Вся геологическая история археоцат (появление, расцвет – около 300 родов, – упадок и вымирание) укладывается в крохотный по геологическим меркам интервал в 15-20 млн. лет – случай совершенно уникальный. Уже во второй половине кембрия эта процветавшая в начале периода группа исчезает «как с белых яблонь дым», и при этом на смену ей не приходит никто – то есть конкурентное вытеснение здесь предположить трудно. Создается впечатление, что археоциаты эксплуатировали некий ресурс, который сперва был в изобилии, а затем стал резко дефицитным. Сопоставляя экологию археоциат (по аналогии с современными губками) и изложенную выше картину вендско-кембрийской биосферной перестройки можно предположить следующее. Группа эта процветала в тот относительно краткий и принципиально неустойчивый период, когда развитие пеллетного транспорта уже обогатило придонные слои кислородом, но еще не сконцентрировало большую часть органики внутри осадка; этот расклад, как полагает А.Г. Пономаренко, должен быть весьма благоприятен для «тонких» фильтраторов. Для того, чтобы эксплуатировать открывшийся ресурс, нет нужды в «квалифицированной рабочей силе» – вполне достаточно макроскопических организмов, даже не являющихся многотканевыми. Однако дальнейшее развитие ситуации с запасанием органики в осадке (что позволяет и сделать ее переработку более равномерной, и улучшить снабжение кислородом придонных слоев) резко ухудшает положение бактерий и пассивных фильтраторов, но зато благоприятствует илоедам. Время археоциат кончается, и наступает время трилобитов (рисунок 25, е-ж)[24] .