Жесткая шерсть, характерна для эрдельтерьера, таксы, фокстерьера, брюссельского гриффона и дратхаара. Она образуется в результате действия доминантного гена Wh (Whitney, 1947; Winge, 1950). Считается, что у всех жесткошерстных пород работает один и тот же ген, хотя предположение о том, что их больше, можно принять как рабочую гипотезу. Есть наблюдения, что ген Wh задерживает рост шерсти, тогда генотип llWh- дает волосы длиннее, чем LLWh-, но короче, чем llwhwh.
Витни предположил, что курчавая или скрученная шерсть ирландского водяного спаниеля наследуется рецессивно по отношению к прямой. Условно, ген обозначен k.
Считалось, что курчавая шерсть ретривера является короткошерстной вариацией скрученной шерсти ирландского водяного спаниеля. Бернс и Фразер (1966) сообщают, что при вязке курчавого ретривера с дратхааром щенки имели такую же курчавую шерсть, как у ретривера. Следовательно, курчавая шерсть, возможно, определяется разными генами у водяного спаниеля и ретривера.
Витни описал особенности некоторых скрещиваний, которые указывают на то, что волнистая шерсть кокер-спаниелей и ряда других пород, возможно, наследуется рецессивно по отношению к прямой. Однако, это не более, чем предположение. Сведений для прямых доказательств недостаточно, поэтому, предполагаемый ген может быть предварительно обозначен, как wa.
Пиллингом была описана линия кокер-спаниеля с необычными кольцами волос на плечах и на боках. Некоторые собаки имели, лишь одно кольцо, которое могло располагаться на голове или шее. Этот признак наследовался рецессивно и ген, его определяющий, обозначен как wo.
Витни описал необычную аномалию кожи (в виде складок) или направления роста волос у щенков бладхаунда. В возрасте 1-7 дней у них появлялись длинные, регулярно расположенные волосы, растущие от шеи до задних конечностей. В семь дней они исчезают, и шерсть по всему телу становится нормальной. Результаты скрещиваний позволяют предположить, что этот признак наследуется рецессивно. Ген, кодирующий его обозначен как rp.
Украшающая шерсть или кайма из длинных волос на конечностях и нижней поверхности хвоста является характерной чертой длинношерстных собак. Степень развития этого признака сильно варьирует. Битнер (1947) сообщает, что кроссы между ирландским водяным спаниелем (с незначительным развитием украшающего волоса на хвосте) и кокером или ирландским сеттером (с обильной оброслостью) давали потомство с обильной оброслостью. Это указывает на доминантность признака, но недостаточно для того, чтобы ответить на вопрос, кодируется ли этот признак одним независимым геном или развивается в результате воздействия модифицирующих полигенов на ген длинношерстности. Последнее наиболее вероятно и может служить примером того, как экспрессия гена длинношерстности может быть модифицирована другими генами.
Витни (1947) отмечает, что у длинношерстных, пятнистых собак черные и белые волосы разной длины. У кокера и английского сеттера черные волосы длиннее, чем белые. Черные волосы также длиннее, чем рыжие. Бернс и Фразер (1966) наблюдали подобное и добавляют, что белые волосы в отдельных случаях могут быть той же длины, что и черные, но мягче по структуре. Рыжие волосы также мягче и шелковистее, чем черные.
Бесшерстность — это явная аномалия. Однако этот признак следует рассмотреть в настоящей работе, т.к. он является определяющим для таких пород, как китайская или мексиканская голые собаки. Эти животные не всегда полностью бесшерстны, а имеют варьирующее количество волос на голове и конечностях. Летард (1930) показал, что эти породы образовались в результате действия доминантного гена Hr, который в гомозиготном состоянии летален. Следовательно, все бесшерстные особи этих пород гетерозиготны по Hr (Hr hr).
Описаны случаи, когда бесшерстность наследовалась рецессивно. Поэтому, не стоит считать, что этот признак всегда кодируется доминантным геном Hr.
Бесшерстные мутанты время от времени встречаются среди нормальных собак и некоторые из них могут выживать в более теплом климате или специально отбираться. Если такие животные имеют древнее происхождение, то могут приобрести ауру респектабельности и быть признанными как порода. Насколько это оправдано является спорным вопросом.
СРАВНИТЕЛЬНАЯ СИМВОЛИКА ГЕНОВ
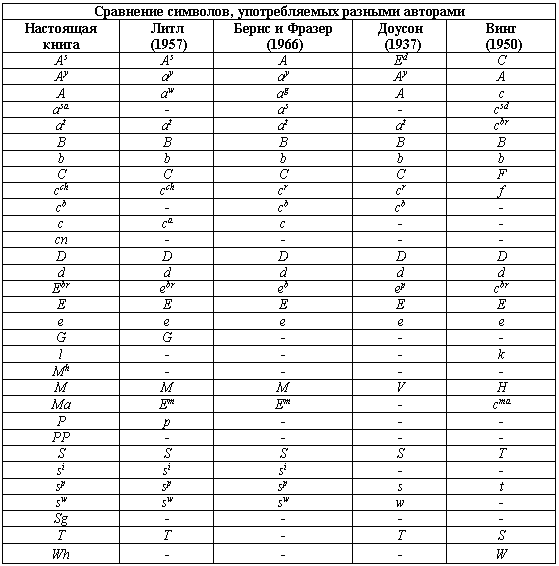
Читатели, которые интересуются литературой по генетике, рано или поздно сталкиваются с проблемой путаницы в обозначениях генов. Дело в том, что различные авторы пользуются различными символами для обозначения одного и того же гена. Это прискорбно, но неизбежно. Исследователи сразу же не согласовали стандартную номенклатуру генов и называли новые описанные гены как им заблагорассудится. Настоящий раздел книги является попыткой свести накопленное в единую систему.
Впервые на это обратил внимание Доусон (1937), который попытался привести в соответствие предыдущие достижения. Собранный им материал выдержал проверку временем. Последующий вклад Литтля (1957), а также Бернса и Фразера (1966) укладывался в основное русло генетики млекопитающих. Увы, это не относится к работам Винга (1951). Его система полностью противоречит предшествующим и достаточно запутана. Он предлагает ряд генов, в которых нет необходимости. Некоторые гены Винг разместил не в тех аллельных сериях, где следовало бы.
Заключение
Если сравнить окрасы собак, обусловленные генами Ay и е, можно заметить, что в первом случае собаки более интенсивно пигментированы. Однако, не стоит считать, что все красные собаки несут Ay, а все более бледные — ее. Там где возникают сомнения по поводу происхождения окраса можно допустить, что он скорее обусловлен Ау, чем ее. Т.е., прежде, чем допустить, что окрас обусловлен генотипом ее, надо доказать, что он не обусловлен геном Ау.
Там, где белая пятнистость является одним из породных признаков, следует учитывать три аллеля, установленных Литтлем (1957). Однако, у многих пород минимальная белая пятнистость является нежелательной или даже запрещенной. Такие отметины возникают чаще всего на морде, копчике хвоста, груди, животе, реже на лапах. Поседение подбородка у многих пород также относится к минимальной белой пятнистости. Сомнительно, чтобы такое многообразие признака кодировались одним геном. Если это так, то этот ген часто изменяет экспрессию самого себя. Это маловероятно, и скорее всего минимальная пятнистость является полигенным признаком, с вариабельной экспрессией. Постоянная выбраковка пятнистых животных означает лишь усиление контроля, но не полная элиминация этого признака. У некоторых пород белые отметины не являются дисквалифицирующим признаком, но нежелательны. Такое послабление понятно, но оно накладывает бремя ответственности, до какой степени допускать проявление этого признака в породе.
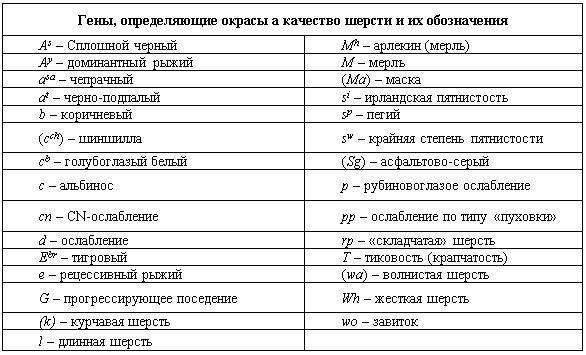
Генетическое обоснование качества шерсти является целой проблемой. К настоящему времени четко установлены лишь несколько генов, определяющих структуру шерсти. Наиболее четко можно идентифицировать гены длинной и короткой шерсти. Несмотря на то, что длинношерстность определяется одним геном l, его экспрессия отличается у многих пород (Витни, 1974).
То же самое нельзя сказать о жесткой, курчавой, волнистой шерсти, шерсти с изломом, с завитком и т.д. Возможно, что у разных пород собак каждый из этих признаков контролируется разными генами. Например, у домашней кошки для шерсти с изломом (порода рекс) идентифицировано несколько генов, хотя фенотипическое проявление их очень сходно. С другой стороны, один и тот же ген в разных породах экспрессируется в разной степени, что может привести к ошибочному мнению о существовании разных аллельных вариантов.