Таблица 6. Размеры конидий из чистой культуры различных географических популяций и моноспорных штаммов
Этот случай лишний раз доказывает, что при изучении внутривидовой или межвидовой изменчивости недостаточно опираться лишь на различия в величине спор изучаемых объектов.
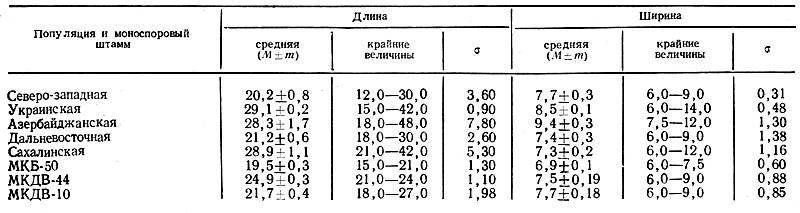
Далее нам интересно было проследить за изменчивостью организма внутри популяции. С этой целью выделенные моноспоровые изоляты также подвергались статистическому анализу. Были взяты биохимические штаммы МКВ-50 (азербайджанский), МКДВ-10 и МКДВ-44 (дальневосточные), которые достоверно отличаются по какому-либо признаку от своих популяций. Здесь также была подмечена закономерность: чем более однороден изучаемый штамм, тем меньше значение сигмы (а), т.е. сигма является достоверным показателем изменчивости ряда. Так, если у азербайджанской популяции а равна 7,8, то у МКБ-50 — 1,3; у дальневосточной популяции — 2,6, у МКДВ-44—1,1.
Таким образом, вид V. inaequalis (Cke) Wint. имеет внутреннюю дифференциацию, что показывает тенденцию вида к дальнейшему развитию и говорит о его жизнеспособности.
Физиологическая специализация гриба. У возбудителя парши яблони имеется ряд физиологических рас, и отдельные сорта яблонь сильно отличаются по своей восприимчивости к различным расам. Развитие эпифитотий зависит от наличия рас, к которым основные сорта яблонь проявляют восприимчивость.
Большой шаг к выяснению взаимодействий хозяина и паразита был сделан генетикой. Ускорилась селекция сортов сельскохозяйственных культур на устойчивость к болезням, и во многих случаях была выявлена генетическая основа устойчивости. Экспериментально было доказано, что виды растений-хозяев обычно содержат огромное количество биотипов и фенотипов и развитие отдельной особи происходит в результате взаимодействий наследственности и условий окружающей среды. Было установлено, что изменения, ранее относимые к влиянию условий окружающей среды, являются генетически контролируемыми.
Кейт, Бун и Шей (1962) в своих исследованиях роли генетических факторов и питания в регулировании взаимодействий хозяина и паразита при парше яблони показали, что парша яблони является удобным объектом для параллельных генетических и биохимических исследований взаимодействия хозяина и паразита. При болезнях, подобных парше яблони, как хозяин, так и паразит могут размножаться половым или бесполым путем, а взаимодействия хозяина и паразита могут изучаться под точным генетическим контролем условий окружающей среды. У возбудителя парши яблони имеется много генотипов с различными уровнями патогенности для разных видов и сортов хозяина. Его можно культивировать на всем протяжении его жизненного цикла и скрещивать in vitro. Изоляты можно размножать бесполым путем с достаточной стабильностью изучаемых признаков, включая патогенность, в течение многих лет. На всем протяжении паразитической фазы и вегетативной стадии in vitro возбудитель находится в гаплоидном состоянии, поэтому можно исследовать эффекты одного набора хромосом.
Сумка содержит все потомство, происходящее от единственного мейоза, с выживанием всех ядер, расположенных в определенном порядке, что позволяет проследить родословную ядерных поколений. Вегетативные клетки одноядерны и могут изучаться независимо от важных проблем гетерокариоза.
Гриб хорошо реагирует на мутагенные воздействия. Кейтом и Буном были получены мутации формы, окраски и биохимического состава. Среди большого количества биохимических мутантов V. inaequalis каждый из 54 изученных признаков наследовался так, как если бы они контролировались одним геном, и почти все случаи дефектов синтеза были полными. Генетическим анализом установлены группы сцепления для шести хромосом. Было найдено, что гаплоидное число хромосом у гриба равно семи.
Кейт и Палмитер (G. W. Keitt, О. Н. Palmiter, 1938) изучали гетероталлизм и изменчивость у V. inaequalis. Полученные восемь моноспоровых штаммов из одной сумки культивировались на листьях яблони. Исследования показали, что не каждые два из этих восьми штаммов могут скрещиваться и давать плодовые тела. Эти восемь штаммов по своим половым особенностям делятся на две группы, и только при скрещивании двух соответствующих штаммов из разных групп можно получить перитеции. Штаммы характеризуются также патогенными особенностями.
По-видимому, устойчивость хозяина зависит от целого ряда генных фондов. На основании реакции специализированных рас гриба (J. R. Shay, E. В. Williams, 1956) хозяев можно разделить на четыре основные группы.
1. Промышленные сорта яблонь, которые полностью восприимчивы в полевых условиях к некоторым линиям патогенных микроорганизмов.
2. Канадский сорт Женева неизвестного происхождения и русские сорта видов Malus baccata и Malus pumila, которые обычно иммунны в полевых условиях, но восприимчивы к определенным расам патогенных организмов.
3. Виды Malus, в основном происходящие из Азии, которые иммунны в полевых условиях и устойчивы ко всем линиям патогенных организмов, насколько это исследовано.
4. Русские и европейские сорта, такие, как Антоновка, которые обладают высокой устойчивостью ко веем исследованным линиям патогенных организмов.
В настоящее время изучается биохимическое регулирование нормальных реакций между линиями дикого типа V. inaequalis и четырьмя группами хозяев.
Кейтт, Бун и Шей (1962) изучали наследование патогенности моноспоровых линий дикого типа V. inaequalis на сортах-хозяевах с фондом генов обычной яблони (Malus sylvestris Mill.).
Опыты показали только два основных типа реакции хозяина к проникновению паразита — «поражение» и «пятно». Реакция «поражение» (восприимчивость хозяина) дает обычные в естественных условиях симптомы, характеризующиеся обильным спорообразованием гриба; реакция «пятно» (устойчивость хозяина) дает хлоротические или некротические пятна с незначительным спорообразованием или без него.
Кейт и Бун (1951, 1957) показали, что патогенность контролируют семь пар локусов. Каждый локус представлен двумя аллелями, одна из которых обусловливает на определенных сортах реакцию типа «поражение», а другая — «пятно».
Приведенные данные свидетельствуют о том, что многообразие и динамичность состава биотипов гриба оказывают глубокое влияние на изменение поражаемости различных сортов, а также и одного сорта в различные годы и в различных эколого-географических условиях.
М. А. Чумаевская (1960) изучала некоторые географические популяции парши яблони и пришла к выводу, подтвердившему высказывания других исследователей, что в условиях Сибири и Алтая имеется особая форма гриба. Сибирские и алтайские популяции гриба способны поражать местные, полукультурные мелкоплодные сорта яблонь — ранетки. Крупноплодные сорта в Сибири парша не поражает или поражает очень слабо.
В Южном Казахстане также имеются, по данным Л. П. Сахаровой (1968, 1971), две резко обособленные расы возбудителя парши, специализированные в отношении дикой и культурной форм яблони. Причем появление парши на культурных сортах яблони отмечается впервые с 1960 г., и связано это явление с интродукцией яблони.
Физиологическая специализация гриба в отношении питающего растения в условиях Ленинградской области изучалась в течение 1946—1948 гг. В. И. Шалышкиной. Исследования показали, что северо-западная популяция гриба широко дифференцирована на многочисленные группы, отличающиеся между собой по морфологическим и культуральным признакам (размерам и обилию конидий, окраске, форме и структуре колоний на искусственных питательных средах).
Физиологическая специализация проявилась в том, что были обнаружены штаммы гриба, изменяющие свои признаки агрессивности в зависимости от сорта.
В отношении штаммов гриба испытывавшиеся сорта разделились на сорта-аккумуляторы и сорта-депрессаторы инфекции. Для первых характерно сильное поражение почти всеми штаммами, для других — слабое поражение. Имеются также сорта пластичные, способные поражаться одними штаммами и не поражаться другими.
Приведенные материалы свидетельствуют о резко выраженной изменчивости гриба V. inaequalis. Эти особенности показывают также, что формообразовательные процессы у возбудителя парши яблони еще более усиливаются в результате сумчатого спороношения и происходящих при этом процессов расщепления признаков и образования новых биотипов в потомстве. Все эти процессы определяют многообразие и динамичность состава биотипов V. inaequalis.
В заключение приводим описание внутривидовых подразделений Venturia inaequalis в пределах СССР.
I группа форм (европейские формы). Размеры аскоспор 14 — 14,3×6,7—7 мкм. Штаммы, относящиеся к этой группе, характеризуются сходными требованиями к температурным условиям. В условиях крайних температур (12 и 32°С) обнаруживают слабый рост мицелия. Оптимальной является температура 22°С.
Северо-западная форма. Отмечена в Ленинградской области. Перитеции имеют средние размеры 234,3×258,4 мкм, аскоспоры — 14,1×6,9 мкм. Размеры конидий с листьев 19,6×6,9 мкм. Конидии из чистой культуры имеют средние размеры 20,2×7,7 мкм. Рост мицелия при температуре 12°С очень слабый, при 32°С почти отсутствует. На добавление к питательной среде дрожжевого автолизата реагирует средне. В культуре гриба преобладает 2-й тип мицелия. На питательных средах спороношение среднее.
Волжская форма. Отмечена в Саратовской области. Размеры перитециев 160,6×160,5 мкм, аскоспор — 14,3×7 мкм, конидий с листа — 19,2×7,4 мкм. Размеры конидий из чистой культуры не представлены, так как штамм очень слабо спороносит на питательных средах. Рост мицелия при температуре 12°С слабый, при температуре 32°С очень слабый. На добавление к питательной среде дрожжевого автолизата реагирует в большой» степени. В культуре гриба преобладает 1-й тип мицелия.