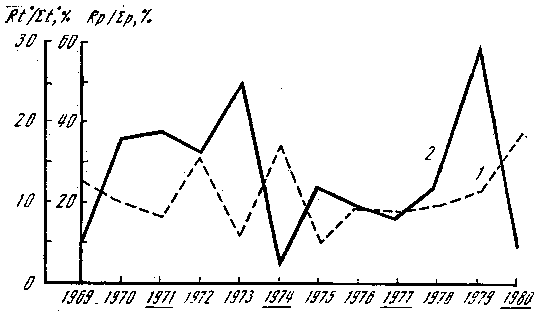
Рис. 8. Количество осадков, суммы положительных температур и плодоношение Armillariella mellea
Rt0 — результирующая температура месяцев (май — август); Rp — результирующая осадков тех же месяцев, мм; Σр — сумма осадков, выпавших к 1 сентября; Σt0 — сумма положительных температур, накопленных к 1 сентября; 1 — Rt0/Σt0, %; 2 — Rp/Σр, %. Подчеркнуты годы массового плодоношения опенка
Рассмотрим вначале зависимость урожайности опенка осеннего (Armillariella mellea) от погодичных изменений температуры. На рис. 8 показана величина, отражающая отношение результирующей накопленных положительных температур четырех месяцев (май — август) к сумме температур за эти месяцы. Она указывает на равномерность накопления тепла; чем ближе к 0° С, тем равномернее распределены температуры в течение исследуемого периода. Абсолютная величина результирующей колеблется от 25 до 320° соответственно в 1979 и 1972 гг. Максимальное количество накопленных положительных температур в июне наблюдалось только в 1969 г., в июле — в 1973, 1974, 1975, 1977 и 1979 гг., в августе — в 1970, 1971, 1972, 1976, 1978 и 1980 гг. Как видно из рисунка, накопление температур происходит с выраженными колебаниями минимума и максимума, период колебаний составляет 2 года. На этом же рисунке отмечены годы с массовым плодоношением опенка. Анализ показывает, что периоды колебаний исследуемых явлений не совпадают. Были рассмотрены и показатели накопленных к 1 сентября (началу плодоношения опенка) положительных температур. Во все исследуемые годы она к этому времени равнялась 2 000—2 500°. Различия эти определяются суммой температур март — конец июня. Месячные и подекадные средние, максимальные и минимальные температуры воздуха вариабельны, а их анализ не дает никаких оснований связывать их величину и изменчивость с периодичностью плодоношения опенка. Можно лишь отметить, что массовому появлению плодовых тел гриба соответствуют величины абсолютного минимума температур — от —1 до +3°, абсолютного максимума — от +18 до +22° в августе.
Как было отмечено, наиболее резкие различия в накоплении суммы положительных температур наблюдаются в весенние месяцы и особенно в апреле. Амплитуда колебаний этой величины по годам достигает 275°, однако высоким урожаям опенка сопутствуют значения суммы положительных температур в очень узких пределах — 80—90°. Это единственная закономерность, которую нам удалось обнаружить.
Количество выпавших осадков в исследуемые годы резко различается в своем распределении по месяцам, декадам и сумме к 1 сентября. Такого четкого и ритмичного колебания минимума и максимума, которое было отмечено для суммы положительных температур, в случае с осадками не наблюдается. Об этом свидетельствуют данные, представленные на рис. 9. Отношение результирующей четырех месяцев (май — август) к сумме осадков, выпавших за эти месяцы, указывает на резко выраженную их неравномерность. Причем никакой зависимости между массовым плодоношением опенка и распределением осадков не наблюдается. Однако можно отметить, что урожайным годам соответствует более или менее равномерное распределение осадков, когда величина отклонения их в различные месяцы не превышает 5—8%. Между абсолютным количеством осадков, выпавших к началу плодоношения опенка, а также ежемесячным и подекадным их режимами и появлением плодовых тел гриба зависимости не найдено. Некоторую информацию о связи этих явлений дает количество осадков, выпавших в последнюю декаду августа, перед непосредственным началом плодоношения. Сумма их колеблется от 3 до 5 в 1971, 1974 и 1977 гг. до 32 мм в 1980 г.
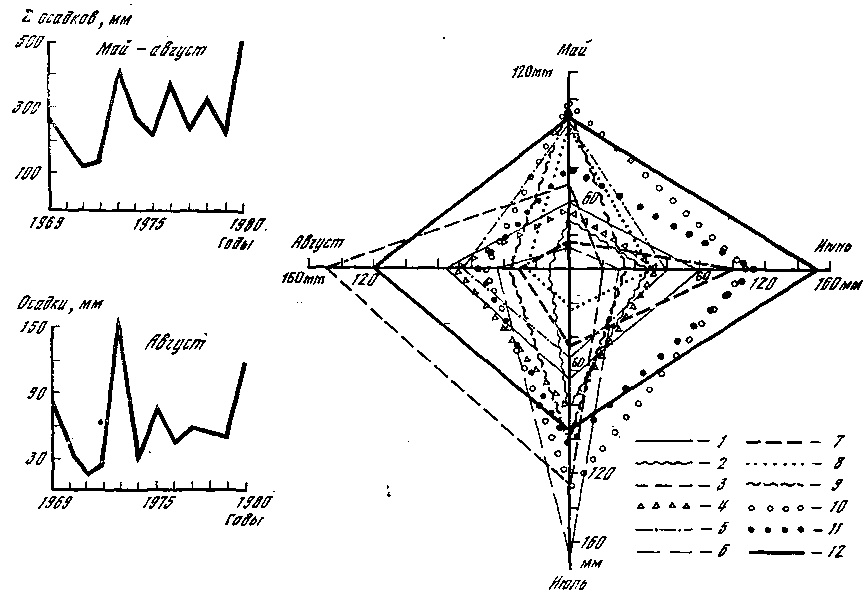
Рис. 9. Амплитуда колебаний выпавших в 1969—1980 гг. осадков
1 — 1969; 2 — 1970; 3 — 1971; 4 — 1972; 5 — 1973; 6 — 1974; 7 — 1975; 8 — 1976; 9 — 1977; 10 — 1978; 11 — 1979; 12 — 1980
Одновременный анализ распределения температур и осадков в связи с цикличностью плодоношения опенка показывает, что урожайные годы характеризуются довольно резкими различиями в показателях суммы накопленных положительных температур и количества выпавших к 1 сентября осадков. В 1971 г. максимальное количество осадков выпало в июле на фоне равномерного накопления температур; 1974 г. (год массового появления опенка) характеризуется жарким июнем при влажном июле; 1977 г. — максимально влажным июнем и жарким июлем и, наконец, 1980 г. — самыми высокими показателями температуры и осадков в августе. Периодичность массового развития опенка, как видно из рис. 9, составляет 3 года и не совпадает с рассмотренными показателями температурного режима и режима влажности. По-видимому, такая цикличность урожаев опенка связана с иными причинами, такими, как, например, физиологические особенности развития гриба, в частности накопление достаточного количества веществ и энергии в мицелии.
Периодичность плодоношения макромицетов зависит от сочетания очень многих факторов, На рис. 10 показана многолетняя динамика количества видов и массы грибов различных экологических групп в одном и том же типе леса. Амплитуда колебаний массы грибов всех групп равна 14,8 кг/га, т. е. приближается к среднемноголетним значениям урожая плодовых тел. Такие различия определяются экстремальными годами, каковым оказался 1972 год. В основном же для лесов Подмосковья различия в урожайности грибов составляют 5—6 кг/га и совпадают по цикличности с суммой накопленных положительных температур за период май — сентябрь. Величина этого цикла — 2 года, а поскольку основу массы грибов составляют макромицеты с крупными плодовыми телами, относящиеся к симбиотрофам, можно считать, что периодичность урожаев таких грибов, как сыроежки, грузди, рядовки, паутинники, белые и березовики, составляют именно 2 года.

Рис. 10. Динамика трофической структуры и биомассы макромицетов (липо-ельник зеленомошно-волосистоосоковый)
Число видов: 1 — симбиотрофов; 2 — ксилотрофов; 3 — гумусовых сапротрофов; 4 — подстилочных сапротрофов; 5 — сапротрофов на опаде; 6 — грибов других трофических групп. Жирной линией обозначена масса макромицетов, кг/га (абс. сухой вес); N — число видов; М — масса грибов
Наибольшей стабильностью в отношении видового состава, численности и массы обладают ксилотрофы и гумусовые сапротрофы, мицелий которых не испытывает в силу своего пространственного положения (защищенности) таких резких колебаний температуры и влажности, как мицелий грибов, расположенных в верхних слоях подстилки. Кроме того, как указывалось ранее, ксилотрофы имеют специфические приспособления для перенесения неблагоприятных погодных условий. Однако самыми консервативными в отношении количества видов и массы оказываются в многолетней динамике сапротрофы на опаде. Объясняется это уникальными физиологическими особенностями плодовых тел этих грибов. На опавших хвоинках и веточках хвойных пород постоянно присутствует в «увядшем» состоянии большое количество хрящеватых тонких (до одного миллиметра) ножек плодовых тел с зачатками шляпок (рис. 10). При повышении относительной влажности приземного слоя воздуха до 70—80% они тут же восстанавливают тургор тканей с минимальной затратой на этот процесс веществ и энергии.
Подстилочные сапротрофы наиболее уязвимы в отношении погодных условий, так как их мицелий, находящийся в различных слоях подстилки, слабо защищен от перепадов температуры и влажности, у них нет симбионтов, которые могли бы помочь им в защите, и нет специальных приспособлений, как у ксилотрофов (ризоморфов), для перенесения неблагоприятных условий.
Масса и количество видов макромицетов в течение одного сезона постепенно увеличиваются с мая, достигая максимальных значений в августе — начале сентября, а затем резко или медленно в зависимости от погодных условий конкретного года уменьшаются. Заканчивается плодоношение всех представителей экологических групп грибов в октябре — начале ноября. Самыми холодостойкими являются некоторые виды ксилотрофов и симбиотрофов, замерзающих на корню (опята, белые, рядовки).