Линн Маргулис назвала внезапные и кардинальные изменения, происходящие вследствие симбиотического объединения геномов вируса и носителя, «эволюционным эквивалентом удара молнии». Но чтобы понять, каков будет эффект этого «удара молнии» в долгосрочной перспективе, и в особенности чтобы понять, каким образом это сыграло ключевую роль в формировании человеческого генома, нам следует изучить подробнее работу естественного отбора для ряда голобионтических партнерств вируса и носителя.
Можно узнать многое о механике подобного генетического союза на примере важнейшего симбиотического партнерства, способствовавшего созданию человеческого генома, — древнего эволюционного переворота, давшего нашим клеткам митохондрии, а нам — возможность дышать кислородом.
Цитируя Льюиса Томаса: «Нас сдают внаем, нас разделили и заняли. Внутри наших клеток живут, даруя энергию, позволяющую нам радоваться солнечному дню и стремиться к совершенству, сотни и тысячи созданий, в строгом смысле нам не принадлежащих, обитающих в цитоплазме — части клетки, лежащей между ядром и оболочкой»[37]. Большинство биологов принимают эволюционное объяснение их существования, впервые выдвинутое Маргулис в конце шестидесятых годов двадцатого века: более миллиарда лет назад дышащие кислородом бактерии вступили в голобионтическое партнерство с одноклеточным протистом (раньше их называли «протозоа»), который и стал предком всех животных, растений, грибов и дышащих кислородом протистов, населяющих сегодня Землю. Снова цитируя Томаса: «Они настолько же симбионты, насколько и ризобиальные бактерии, живущие в корневой системе бобовых и позволяющие накапливать азот. Без митохондрий мы бы не могли двинуть ни единым мускулом, не могли бы согнуть палец и думать не могли бы».
Чем же можно доказать подобное утверждение?
Доказательств известно несколько. Геном митохондрий составлен из типичных бактериальных генов, и они, в отличие от наших ядерных генов, соединены в кольцо, что характерно для бактерий. Размножаются митохондрии независимо от ядра, типичным для бактерий процессом почкования. Геномы митохондрий всех современных имеющих митохондрии форм жизни настолько генетически схожи, что они, несомненно, произошли от общего предка. Как ни удивительно, генетический анализ показывает: ближайший из современных родственников предка всех митохондрий — это бактерия Rickettsia prowazeki, возбудитель печально известного заболевания — разносимого вшами эпидемического тифа. Теперь нам известно: этот возбудитель тифа не может существовать самостоятельно, он чрезвычайно интенсивно (и зачастую агрессивно) взаимодействует с клетками носителя, геном его естественным отбором редуцирован и рудиментарен и не позволяет бактерии жить вне клеток-носителей.
Если изучить, как естественный отбор влиял на геном наших митохондрий в течение длящегося миллиард лет голобионтического симбиоза с ядерными клетками, обнаружится, что геном предка митохондрий претерпел радикальные изменения — был разрушен, упрощен. Большое число изначально присутствовавших генов исчезло, было выброшено за ненадобностью, а около трехсот генов переместились от митохондрии в ядро, где, в подобии уже обсуждавшегося здесь сожительства Elysia chlorotica с чужими хлоропластами, эти перенесенные гены продолжают работать согласованно с остатком генов митохондрии. Но не может не удивлять то, что за миллиард лет симбиоза геном митохондрии сохранил отчетливо бактериальный характер. Как мы увидим далее, понимание симбиотического происхождения человеческих митохондрий важно для изучения заболеваний, обусловленных нарушениями митохондриального генома.
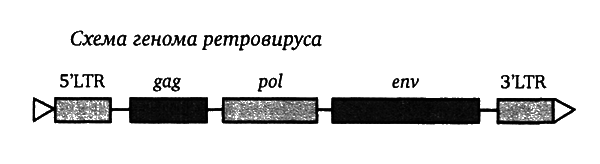
Приведенный выше рисунок иллюстрирует структуру генома типичного ретровируса — такого, как вызывающий СПИД ВИЧ-1, или KoRV, вызывающий лейкемию у коал, или любой из 30–50 семейств HERV, входящих в обширную коллекцию вирусных компонентов человеческого генома.
При взгляде на этот рисунок видно: геном начинается и кончается с областей, обозначенных как «LTR» — это сокращение английского выражения «длинный концевой повтор» (long terminal repeat), и эти области — не гены как таковые, но «бюрократы» с чрезвычайно мощными «управленческими» способностями. Такие «бюрократы» встречаются только у вирусов, именно они несут способность манипулировать генетическим аппаратом носителя. Собственно вирусные гены, управляющие жизненным циклом ретровируса, содержатся в трех областях, обозначенных как gag, pol и env (эти обозначения принято писать курсивом). В области gag содержатся гены, кодирующие белки матрицы вируса (специфичные антигены), область pol кодирует важнейший для ретровируса энзим, обратную транскриптазу, и другие важные энзимы, включая «интегразу», позволяющую вирусной ДНК залезть в хозяйскую хромосому, область же env кодирует белки оболочки вируса.
С традиционной дарвиновской точки зрения, любое вторжение вируса в геном хозяина можно интерпретировать как «эгоистическое» действие с целью выживания этого вируса, и, вполне возможно, естественный отбор будет действовать во многом индивидуально для вируса и носителя. Однако, как мы уже видели на примере СПИДа, даже в разгар жесточайшей эпидемии можно наблюдать и отбор, работающий в значительной мере на уровне симбиоза. Во время активного распространения эпидемии логично ожидать неприятных последствий агрессивного симбиоза и «выбраковки» значительной части вида-носителя. Но, судя по эпидемии у коал, эта стадия может длиться не так уж много времени. «Выбраковка» приводит к созданию нового партнерства между вирусом и носителем, остатки вида размножаются снова, и эти остатки, активно взаимодействующие с вирусом, будут иметь эволюционное преимущество. Если экзогенная инфекция летальна для носителя, то совокупление носителя эндогенного вируса с теми, кто не носит эндогенный вирус, приведет к их гибели. А носители эндогенного вируса останутся заразными в течение очень долгого времени. Вирусные же элементы в геноме размножатся, внедрив себя в хромосомы, предоставив геному носителя уже готовые гены и мощные средства управления ими, — и это способно радикально изменить эволюционную траекторию новой сущности, нового голобионтического организма.
Вполне логично ожидать, что за сотню миллионов лет голобионтического партнерства млекопитающих и вирусов естественный отбор сделает с эндогенными вирусами то же, что и с митохондриями: выбросит ненужные вирусные гены и «бюрократические» управляющие элементы — но вирусов в геноме так много, что некое активное меньшинство будет присутствовать всегда. Оно всегда будет играть важную роль в голобионтической эволюции. Следует также ожидать, что вирусы даже после внедрения в наш геном и превращения в его часть по-прежнему останутся в сути своей вирусами. Если принять во внимание, что такое голобионтическое партнерство и естественный отбор у человека и его предков происходили с сотнями вирусов (в процессе чего и был создан человеческий геном), то понятной становится структура произошедших от вирусов частей человеческого генома.
Вильярреал полагает, что LINE и SINE, вместе составляющие тридцать четыре процента человеческого генома, либо являются производными ретровирусов, либо контролируются ими. Вывод обескураживающий и поразительный. Это значит: вирусы вместе с их производными и тесно связанными структурами составляют как минимум сорок три процента человеческого генома — и со временем, по мере дальнейшего исследования еще не расшифрованных частей генома, этот процент, скорее всего, увеличится. Скептики вполне резонно могут потребовать доказательств. И как же доказать столь созидательную роль симбиоза с вирусами в происхождении нашего генома? Для этого нужно искать доказательства естественного отбора на голобионтическом уровне при взаимодействии вируса с носителем. Более того, нужно продемонстрировать и важность геномных ретровирусов и их производных в человеческой генетике, развитии эмбриона, физиологии — и, конечно же, в возникновении человеческих заболеваний.
37
Thomas L. The Lives of a Cell. London: Penguin Books, 1974: 4.