
Рис. 17. Репродуцирование разрывом
Репродукция клетки
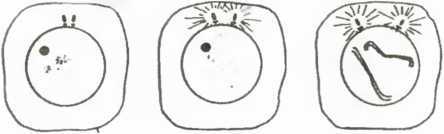
Какое отношение все это имеет к клеткам? Если взять любую клетку на стадии интерфазы (т. е. не во время репродуктивного процесса) и «разломать» ее, то двух клеток мы не получим. На стадии интерфазы клетка представляет собой компартменталиэованную систему, т. е. существуют компоненты, изолированные от остальных или представленные в единственном числе (что исключает наличие любой плоскости репродуктивного разделения). В частности, это справедливо для молекул дезоксирибонуклеиновой кислоты (ДНК), образующих составную часть хромосом и отделенных на стадии интерфазы ядерной мембраной от цитоплазмы в ядре (рис. 18 а).
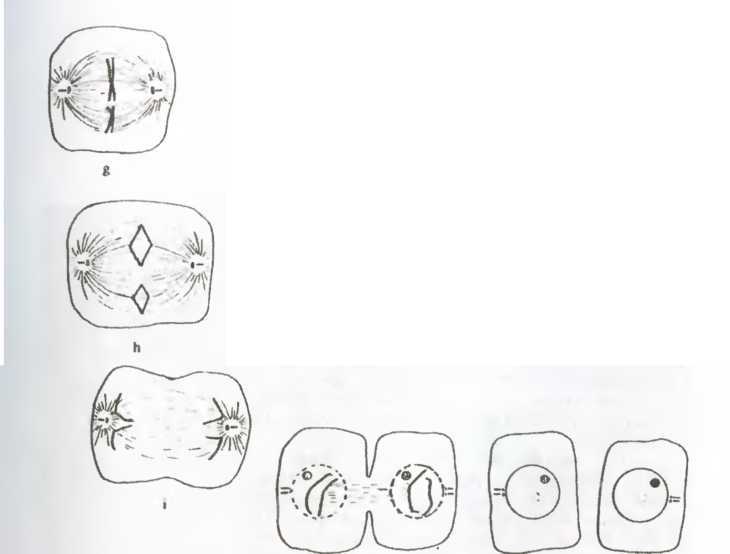
При митозе, или клеточном делении, все процессы (рис. 18 b-j) сводятся к декомпартментализации клетки. Это отчетливо видно на рис. 18, где показано растворение ядерной мембраны (сопровождающееся репликацией больших молекул ДНК, имеющих вид двойных спиралей) и смещение хромосом и других клеточных компонентов, что делает возможным появление плоскости раздела. Все это происходит в результате клеточного аутопоэза, который при этом не прерывается. Та ким образом, собственная динамика клетки ведет к структурным изменениям, например, к образованию митотического веретена (рис. 18 d-h). Такие изменения приводят к дроблению, или делению клетки.
С этой точки зрения процесс репродукции клетки достаточно прост и сводится к «разлому» вдоль плоскости деления,
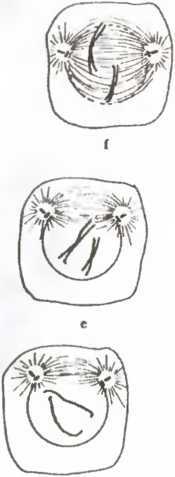
что порождает два новых единства того же класса, что и исходная клетка. В современных эукариотных (имеющих ядро) клетках плоскость деления и механизм деления определяются тонким и изящным механизмом молекулярной хореографии. В древних прокариотных клетках, где компартментализация, изображенная на рис. 18, не наблюдается, процесс репродукции в действительности происходит проще. В любом случае репродукция клетки — это именно репродукция в указанном выше смысле, а не репликация или копирование единств.
Но в отличие от приведенных выше примеров репродукции, деление клетки — явление специфическое: оно обусловлено аутопоэзной динамикой. Никакие внешние агенты или силы для него не требуются. Мы можем лишь предполагать, что в случае первых ау-топоэзных единств дело обстояло иначе и что в действительности репродукция сначала была фрагментацией, возникавшей при столкновениях этих единств с другими внешними объектами. В получавшейся при этом исторической сети некоторые необычные клетки подвергались репродуктивному делению вследствие своей внутренней динамики. Такие клетки обладали механизмом деления, положившим начало наследственному ряду, или устойчивой исторической последовательности. Как именно это произошло, остается неясным. Возможно, мы никогда этого не узнаем. Но это отнюдь не обесценивает тот факт, что клеточное деление представляет собой частный случай репродукции, который мы на вполне законном основании можем назвать саморепродукцией.
Рис 18 Митоз, или репродуцирование разрывом, в клетке животного. На рисунке изображены различные стадии декомпартмен-тализации, делающей возможной репродуцирование разрывом
Репродуктивная наследственность
Всякий раз, когда мы имеем дело с историческим рядом, независимо от того, как он возник, мы сталкиваемся с феноменом нвследственности, т. е. обнаруживаем, что структурные конфигурации, присущие какому-то члену ряда, вновь появляются у следующего члена ряда. Это справедливо как в отношении характеристик, присущих всему классу, так и для признаков отдельных особей. Поразмыслив над примером с историческим рядом последовательных ксероксных копий, мы обнаружим, что сколь бы сильно первые копии ни отличались от последних, определенные соотношения между черным и белым цветом в начертаниях букв останутся неизменными; именно это и позволяет нам читать копии и утверждать, что одна страница является копией другой. В тот момент, когда копия станет настолько неразборчивой, что мы не сможем прочитать ее, исторический ряд прервется
Аналогичным образом в системах, наделенных способностью репродуцирования, наследственность присутствует в каждом репродуктивном акте как составная часть феномена репродукции в силу одного лишь факта порождения двух единств одного и того же класса. Иначе говоря, именно потому, что репродукция происходит, когда существует плоскость деления в единстве с распределенной структурой, при переходе от одного поколения к следующему неизбежно сохраняется определенная устойчивость структурных конфигураций.
Точно так же, поскольку репродуктивное деление приводит к образованию двух единств с той же организацией, что у исходного единства, но с отличающимися структурами, оно сохраняет организацию и порождает структурные вариации. Иэ феномена репродукции с необходимостью следует как возникновение сходства, так и структурных различий между «родителями», «детьми» и «братьями» Те аспекты первичной структуры нового единства, которые мы оцениваем как идентичные исходному единству, и называются наследственностью; те же аспекты первичной структуры нового единства, которые мы оцениваем как отличные от исходного единства, называются репродуктивной вариацией Именно поэтому каждое новое единство неизменно начинает свою индивидуальную историю, обладая и структурным сходством со своими предками, и отличиями от них. Как мы увидим, эти сходства и различия сохранятся или будут утрачены в зависимости от обстоятельств в процессе конкретного онтогенеза Теперь же мы хотим лишь подчеркнуть, что репродукции присущи и феномен наследственности, и появление структурных изменений у потомков; это утверждение остается в силе и относительно репродукции живых существ.
В репродукции клеток имеется немало примеров, когда удается точно установить структурные условия, обусловливающие как появление изменений, так и сохранение сходства. Например, некоторые компоненты допускают лишь ограниченное число вариантов своего участия в аутопоэзе, но многочисленные режимы своего участия в клеточной динамике. Такие компоненты входят в основные структурные конфигурации, переходящие из поколения в поколение (в противном случае репродукции бы не было) почти без изменений. служить ДНК (нуклеиновые кислоты) или гены, основная структура которых при репродукции воспроизводится лишь с небольшими изменениями, что приводит к сохранению основных структурных конфигураций у особей одного наследственного ряда. В то же время, существует непрерывная изменчивость структурных аспектов, которая не позволяет сохранять их более чем в одном или двух поколениях. Например, способ синтеза белков с участием ДНК сохранялся неизменным во многих наследственных рядах, тогда как тип синтезируемых белков на протяжении истории этих наследственных рядов претерпел глубокие изменения.
Характер распределения структурных изменений или инвариантности в системе наследственных рядов определяет различные способы проявления наследуемости в череде поколений, иными словами — различные генетические (наследственные) системы. Современные генетические исследования сосредоточены главным образом на генетике нуклеиновых кислот. Но существуют и другие генетические (наследственные) системы, которые мы только начинаем понимать. Они остались в тени генетики нуклеиновых кислот — к примеру, генетика других внутриклеточных образований, например, митохондрий и мембран.
4. Жизнь метаклеточных
Под онтогенезом мы понимаем историю структурных изменений, происходящих в некотором единстве без потери этим единством своей организации. Такие структурные изменения, вызываемые либо взаимодействиями единства с окружающей средой, в которой оно существует, либо его внутренней динамикой, происходят непрерывно. Что касается постоянных взаимодействий с окружающей средой, то клеточное единство ежемоментно классифицирует и «рассматривает» их в соответствии со своей структурой. В свою очередь, структура единства непрерывно изменяется вследствие его внутренней динамики. В итоге онтогенетическое преобразование единства прекращается только с его распадом. Для того, чтобы кратко представить эту ситуацию, говоря об аутопоэз-ных единствах, мы будем использовать следующую диаграмму: