Эта сравнительная простота, казалось бы, говорит о том, что гладкие мышцы представляют собой один из этапов эволюции скелетных мышц. Гладкая мускулатура тоже способна к сокращению и, несмотря на отсутствие сложной микроскопической структуры, работает совсем неплохо. Однако исследования мышечных белков различных видов животных показывают, что эволюция мускулатуры была вовсе не так проста. Одно тщательное исследование, которое провели генетики Оота Сатоси и Сайтоу Наруя из японского Национального института генетики в Мисиме, показало, что набор белков, действующих в скелетных мышцах млекопитающих, так похож на набор белков в поперечнополосатых летательных мышцах насекомых, что они явно унаследованы от общего предка позвоночных и беспозвоночных, жившего около шестисот миллионов лет назад. У этого существа уже были поперечнополосатые мышцы, хотя и не было скелета. То же самое относится и к белкам гладких мышц, эволюция которых восходит к другому давнему общему предку. Гладкая мускулатура не была промежуточным этапом на пути к более сложной поперечнополосатой мускулатуре — это просто другая эволюционная ветвь.
Это весьма примечательный факт. Миозин наших собственных скелетных мышц состоит в более близком родстве с миозином, за счет которого летает назойливая комнатная муха, которая вьется у вас над головой, чем с миозином из ваших собственных сфинктеров, сжимающихся от отвращения. Как ни удивительно, они разделились даже задолго до нашего общего предка и, судя по всему, еще до возникновения билатеральной (двусторонней) симметрии, свойственной как насекомым, так и позвоночным. Похоже, что даже у медуз есть поперечнополосатые мышцы, во многих деталях похожие на наши. Итак, и гладкие, и поперечнополосатые мышцы сокращаются с помощью похожей системы из актина и миозина, но те и другие, судя по всему, развились независимо и унаследованы нами от общего предка, у которого были оба типа мышечных клеток. Причем этот общий предок был одним из древнейших животных, существовавших в те времена, когда венцом творения были какие-нибудь медузы.
И все же, несмотря на неожиданно продолжительное эволюционное разделение поперечнополосатой и гладкой мускулатуры, ясно, что все бесчисленные формы миозина восходят к одной предковой форме. Все они обладают одной и той же базовой структурой, все они связываются с актином и АТФ в одних и тех же местах, и все они проворачивают один и тот же двигательный цикл. Если миозины поперечнополосатой и гладкой мускулатуры происходят от общего предка, значит, этот предок был еще примитивнее медуз, и у него, по-видимому, не было ни поперечнополосатых, ни гладких мышц. И все же он находил какое-то применение и актину, и миозину. Какое? Ответ на этот вопрос был получен еще в 60-х годах XX века — благодаря неожиданному открытию. Но хотя это открытие было сделано уже довольно давно, его по праву можно считать одним из самых поразительных в истории биологии. Оно удивляет не только своей наглядностью, но и тем, сколько света оно пролило на эволюцию мышц. Совершил это открытие Хью Хаксли. Он обнаружил, что актин можно «украсить» миозиновыми головками и рассматривать в электронный микроскоп. Сейчас я все объясню.
Актиновые и миозиновые нити можно извлечь из мышечных волокон и разделить на составляющие. Например, головки миозина можно отделить от их длинных хвостов и соединить в пробирке с актином. Актин может сам быстро собираться в длинные нити. Способность к полимеризации при соответствующих условиях — одно из его неотъемлемых качеств. К таким нитям миозиновые головки тоже прикрепляются сами, как и в работающих мышцах, выстраиваясь вдоль актиновых нитей как стрелки, которые рисуют на схемах. Все эти стрелки указывают одно направление, отражающее полярность актиновых нитей, всегда собирающихся в одной и той же конфигурации. Миозин тоже всегда связывается с ними в одном и том же направлении, что позволяет создавать мышечную силу. (В саркомерах актиновые волокна, расходящиеся в стороны от середины, имеют противоположную полярность, так что при сокращении оба края саркомера приближаются к середине и весь саркомер сжимается. Такое сокращение множества расположенных в ряд саркомеров приводит к укорачиванию всей мышцы.)
Миозиновые головки связываются только с актином и ни с чем иным, поэтому, добавляя их к содержимому любых клеток, можно проверять, есть ли в них актиновые нити. До 60-х годов считалось, что актин — это специализированный мышечный белок, неизменно присутствующий в мышцах разных видов животных, но отсутствующий в их клетках других типов. Эти общепринятые представления лишь недавно были поставлены под сомнение биохимическими данными, указывавшими на то, что у одного из многих организмов, не имеющих мышц, — у пивных дрожжей, — актин, возможно, все-таки есть, но когда стало известно, что актин можно украшать миозиновыми головками, открылся настоящий ящик Пандоры. Первым его открыл Хаксли. Он добавил кроличий миозин к актиновым нитям, выделенным из слизевика — представителя очень примитивной группы организмов, — и обнаружил, что эти белки прекрасно подходят друг к другу.
Актин есть везде. У всех сложных клеток имеется внутренний скелет (цитоскелет) из актиновых (и других) нитей. Во всех клетках нашего организма, как и во всех клетках любых других животных, а также любых растений, грибов, водорослей и простейших, есть актиновый цитоскелет. И то, что кроличий миозин связывается с актином слизевика, заставляет предположить, что актиновые нити из принципиально разных типов клеток очень похожи по своей тонкой структуре. Это предположение, как ни удивительно, совершенно справедливо: например, теперь нам известно, что генетические последовательности, кодирующие актин у человека и у дрожжей, совпадают на 95 %[48]. В свете этого эволюция мышц выглядит совсем по-другому. Те же самые нити, которые работают в наших мышцах, работают и во всех сложных клетках. Они по-настоящему отличаются только тем, что по-разному организованы.
Среди всех музыкальных форм мне почему-то особенно дороги вариации. Утверждают, что когда Бетховен в юности демонстрировал свою игру Моцарту, она не произвела на Моцарта особого впечатления, если не считать мастерства импровизации — умения извлекать бесконечные ритмические и мелодические вариации из одной простой темы. В зрелые годы это мастерство достигло у Бетховена апогея в его великих Вариациях на тему Диабелли. Как и предшествовавшие им замечательные баховские Вариации Гольдберга, бетховенские вариации отличаются строгостью формы. Положенная в их основу гармоническая схема сохраняется на протяжении всего произведения, сообщая ему безошибочно ощущаемое единство. В последующие времена композиторы часто отказывались от подобной строгости, что позволяло им задерживаться на различных настроениях и впечатлениях, но лишало их произведения ореола математического совершенства. Такие вариации не создают ощущения, что все скрытые нюансы мелодии в них извлечены на свет, все тайные измерения обрели реальность, все возможности были использованы.
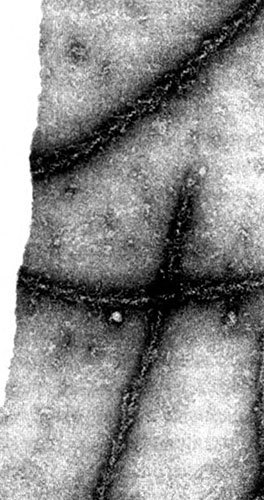
Рис. 6.3. Актиновые нити, добытые из слизевика Physarum polycephalym, украшенные актиновыми «стрелками» из мышц кролика.
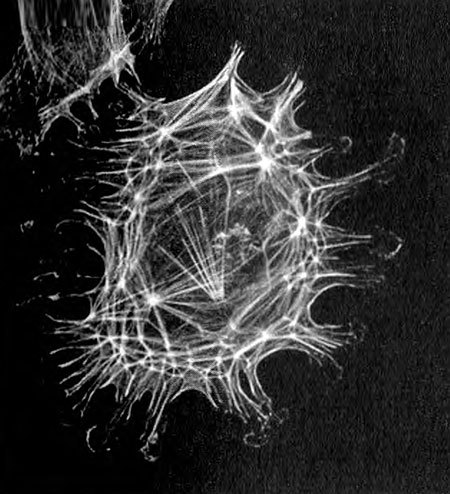
Рис. 6.4. Актиновый цитоскелет в клетке хрящевой ткани коровы, помеченный флуоресцентным красителем фаллоидином-ФИТЦ.
Эта способность взять определенную тему и проиграть ее во всевозможных вариациях, неизменно сохраняя верность основным элементам ее структуры, характерна и для живой природы. Например, на такую заданную тему, как двигательные взаимодействия миозина и актина, естественный отбор с бесконечной изобретательностью писал вариации, породив захватывающий дух набор форм и функций. Внутренний мир любой сложной клетки наглядно подтверждает, с какой удивительной легкостью естественному отбору даются строгие вариации.
48
На самом деле ситуация несколько сложнее: сами генетические последовательности совпадают на 80 %, а на 95 % совпадают последовательности аминокислот в кодируемых ими белках. Это возможно потому, что многие аминокислоты могут быть закодированы по-разному (см. главу 2). Разница между приведенными выше показателями сходства — результат обычных мутаций в последовательности ДНК-букв в сочетании с сильным отбором на сохранение исходной последовательности аминокислот. Отбор почти всегда допускает лишь те изменения генетической последовательности, которые не приводят к изменению последовательности аминокислот в белке. Это еще один небольшой, но наглядный пример действия отбора.