Караси-амазонки, исследованные советскими генетиками К. А. Головинской, Д. Д. Ромашовым и Н. Б. Черфас, не представляют собой исключения. Такой механизм известен у живородящей рыбки молинезии из мелких водоемов Центральной Америки. Гиногенетичными оказались многие насекомые, некоторые черви и амфибии. Часть описанных случаев отличается поразительной сложностью.
Один из видов американской саламандры — амбистома латерале (близкий вид — тигровая хорошо известен любителям живой природы под названием аксолотля) в одних и тех же водоемах может быть представлен диплоидной и триплоидной расой. Триплоидные амазонки, более крупные и сильные, чем обычная диплоидная форма, могли бы ее вытеснить и вымереть вслед за ней сами, так как их икринки стимулируются к развитию диплоидными самцами. Однако баланс между формами поддерживается тем, что у триплоидов плодовитость значительно меньше, да и развитие икринки затягивается на более длительный срок.
В последнее время снова возник спор о роли полиплоидизации в эволюции животных, особенно тех, у которых пол не однозначно определяется хромосомным набором, а зависит от соотношения в организме женских и мужских половых гормонов. Особенно стремятся доказать ее широкую распространенность в природе сторонники так называемого «мгновенного видообразования», когда в течение одного поколения возникает новый вид, не смешивающийся с материнской формой. Но на пути подобных построений неизбежно встает парадокс Каина. Растения обходят его, так как способны к самоопылению и вегетативному размножению. Но как быть с животными?
И все же есть факты, свидетельствующие о возможном удвоении генома у двуполых животных. Рыжий таракан-пруссак имеет 24 хромосомы, а крупный черный — 48. У карпа 104 хромосомы, а у большинства других карповых — 52. Лососевые рыбы, как это установлено достаточно твердо, — тетраплоиды. Однако многие подобные случаи оказались проявлением так называемой ложной полиплоидии, обусловленной расщеплением и слиянием хромосом и рядом других факторов. В опыте полиплоидия животных возможна. Непрямым путем — через партеногенез и межвидовую гибридизацию Б. Л. Астаурову и его сотрудникам удалось сконструировать аллотетраплоид — гибрид домашнего и дикого тутового шелкопряда. Я умышленно говорю — сконструировать, потому что, когда читаешь описание этого замечательного опыта, именно это слово приходит на ум. Аллотетраплоид Астаурова, как и рафанобрассика Карпеченко — форма, генетически изолированная от обоих родительских видов. Для того чтобы ее получить, потребовалось:
1) научиться получать партеногенетическое потомство с диплоидным набором хромосом (здесь индуктором развития яйцеклетки выступает не сперма другого вида, а нагрев до 48 °C в течение 18 минут);
2) получить тетраплоидных самок, размножающихся партеногенетически (число хромосом 4n = 112);
3) скрещиванием тетраплоидных самок с обычными диплоидными самцами получить триплоидные формы шелкопряда;
4) из триплоидных форм получить гексаплоидные, с шестью хромосомными наборами;
5) гексаплоидных самок домашнего шелкопряда скрестить с диплоидными самцами дикого. Потомство будет иметь соответственно — три набора хромосом от домашнего и один — от дикого шелкопряда, то есть будет аллотетраплоидным;
6) путем отбора из поколения в поколение повысить плодовитость гибридов.
Значение подобных опытов трудно переоценить, ибо они закладывают фундамент новой отрасли селекции и генетики — получению плодовитых гибридов между разными видами животных. Однако вряд ли виды часто возникали в природе подобным путем, так как арсенал средств природы несравненно беднее, чем у современного экспериментатора. У природы другой козырь — время, в течение которого самые радикальные перестройки генома могут стать в принципе не невозможными…
Теперь остановимся коротко на менее решительных перестройках генома — частичном удвоении числа хромосом, соединении двух хромосом в одну или, наоборот, разделении их. Такие случаи встречаются гораздо чаще, чем авто- и аллополиплоидия.
Долгое время среди генетиков бытовало убеждение, что число хромосом — абсолютно стабильный видовой показатель. Однако оно оказалось не совсем верным. Описаны случаи, когда число хромосом изменяется или путем распада одной хромосомы на две, или путем слияния двух негомологичных (не образующих пары при мейозе) в одну. Возможна и полисомия — удвоение не всего хромосомного набора, а лишь части хромосом. Такие межхромосомные перестройки весьма важны для эволюционного процесса. Увеличение числа хромосом приводит к резкому увеличению комбинаторной изменчивости. Наоборот, слияние их снижает изменчивость, следствием чего является замедление образования форм, стабилизация вида.
В последние годы широко распространено мнение, что первичные, примитивные виды имеют геном, состоящий из многих мелких хромосом, а эволюционный процесс идет путем слияния их в крупные. Возможно, во многих случаях это так — и все-таки мне кажется, что здесь уместно вспомнить гетевское — «теория мертва, лишь древо жизни вечно зеленеет». Эволюцию нельзя загонять в узкие рамки одной тенденции. Есть прямые свидетельства того, что изменения генома шли в разных направлениях, многократно комбинируясь. Примером могут служить олени. До наших дней сохранились мелкие примитивные тропические оленьки — мунтжаки; в той же группе есть и значительно эволюционно подвинутые виды. Оказывается, у большинства видов оленей 50–70 хромосом, у примитивного оленя Давида, как у лани и благородного оленя, 2n = 62, у одного из видов мунтжаков (мунтжак Ривса) — 46, а у другого — только 6, как у комара, наименьшее известное для млекопитающих хромосомное число! Комментарии, как говорится, излишни.
Менее заметны при цитологическом анализе, но довольно часто встречаются такие хромосомные перестройки, как инверсии и транслокации. В случае инверсии участок хромосомы перевертывается на 180°; такая перестройка затрагивает только одну хромосому. При транслокации происходит обмен участками между двумя негомологичными хромосомами (отличие от кроссинговера!).
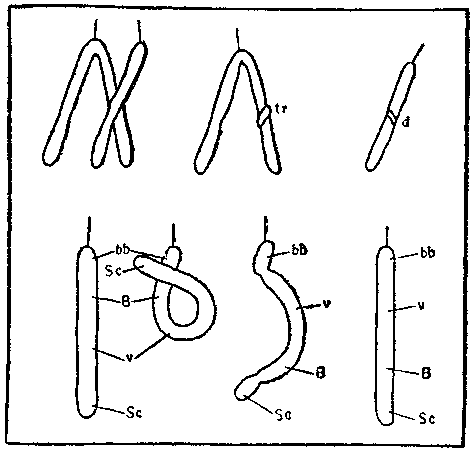
Единая схема возникновения мутации (по А. С. Серебровскому, 1929). Верхний ряд — образование транслокаций (tr) и нехваток (d); нижний ряд — образование инверсии; Sc, v, В, bb — символы разных генов.
Замечательный советский генетик Александр Сергеевич Серебровский еще в 1929 году расшифровал механизм возникновения инверсий и транслокаций — разрыв хромосомы с последующим воссоединением в другом месте. Если хромосома разорвется сразу в двух точках, оторванный участок может, перевернувшись на 180°, воссоединиться с остальным телом хромосомы. Разорванные концы хромосом, как оказалось, «слипаются» с высокой вероятностью (молекулярный механизм этого явления мы начинаем угадывать только сейчас).
Но если разрыв произошел в одном месте, «поворота кругом» не происходит. Конец хромосомы, именуемый теломером, воссоединяться даже со свежим разрывом не может, в данном случае возможно лишь восстановление прежней структуры. Если же она не восстанавливается, оторванный кусок бесследно рассасывается, и генетическая информация, заключенная в нем, пропадает. Такие аварии называют концевыми нехватками или дефишенси. Судьба организма, геном которого потерпел дефишенси, складывается по-разному. Если потерянный кусок мал и не содержит жизненно важных генов, организм, особенно полиплоидный, это легко перенесет. Потеря большого куска хромосомы может быть смертельной. Например, потеря значительной части одной хромосомы в геноме человека приводит обычно к смертельной генетической болезни, называемой «синдромом мяукания» (cri de chat — по звукам, издаваемым больными младенцами).
В случае двойного разрыва хромосомы оторванный кусок также может рассосаться в плазме (воссоединяется только концевой кусок с теломером). Такие нехватки называют делециями. Описано много случаев делеций у самых различных организмов. Крупные делеции, как правило, в гомозиготном состоянии летальны, мелкие — вызывают самые разнообразные изменения признаков фенотипа.