
В 1949–1952 гг. А. Ходжкин, Э. Хаксли, Б. Катц создали со-временную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30 -50 раз больше ионов калия, в 8–10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Проницаемость мембраны для ионов обусловлена ионными каналами, макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, другие (потенциалозависимые) открываются и закрываются в ответ на изменения МП. Потенциалозависимые каналы подразделяются на натриевые, калиевые, кальциевые и хлорные. В состоянии физиологического покоя мембрана нервных клеток в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Таким образом, согласно обновленной мембранной теории асимметричное распределение ионов по обе стороны мембраны и связанное с этим создание и поддержание мембранного потенциала обусловлено как избирательной проницаемостью мембраны для различных ионов, так и их концентрацией по обе стороны от мембраны, а более точно величину мембранного потенциала можно рассчитать по формуле.
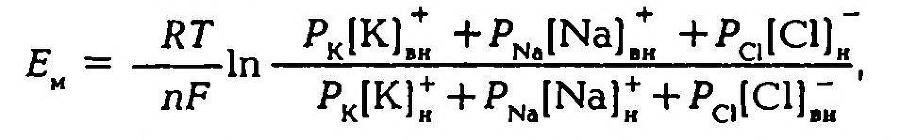
Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т. е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы – крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд. Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки. То что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса.
Изменения мембранного потенциала. Пороговые и подпороговые раздражители
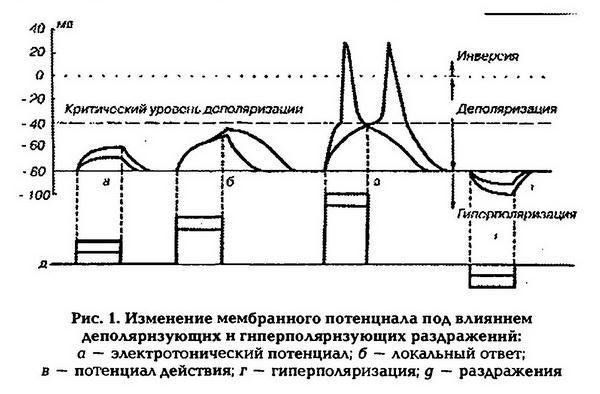
Если раздражать нейрон через электрод, находящийся в цитоплазме, кратковременными импульсами деполяризующего электрического тока различной величины, то, регистрируя через другой электрод изменения мембранного потенциала, можно наблюдать следующие биоэлектрические реакции: электротонический потенциал, локальный ответ и потенциал действия (рис.1). Если наносятся раздражения, величина которых не превышает 0,5 величины порогового раздражения, то деполяризация мембраны наблюдается только во время действия раздражителя. Это пассивная электротоническая деполяризация (электротонический потенциал). Развитие и исчезновение электротонического потенциала происходит по экспоненте и определяется параметрами раздражающего тока, а также свойствами мембраны (ее сопротивлением и емкостью). Во время развития электротонического потенциала проницаемость мембраны для ионов практически не изменяется.
Локальный ответ.
При увеличении амплитуды подпороговых раздражений от 0,5 до 0,9 пороговой величины развитие деполяризации мембраны происходит не прямолинейно, а по S-образной кривой. Деполяризация продолжает нарастать и после прекращения раздражения, а затем сравнительно медленно исчезает. Этот процесс получил название локального ответа.
Локальный ответ имеет следующие свойства:
1. возникает при действии подпороговых раздражителей;
2. находится в градуальной зависимости от силы стимула (не подчиняется закону «все или ничего»); локализуется в месте действия раздражителя и не способен к распространению на большие расстояния;
3. может распространяться лишь локально, при этом его амплитуда быстро уменьшается;
4. локальные ответы способны суммироваться, что приводит к увеличению деполяризации мембраны. В период развития локального ответа возрастает поток ионов натрия в клетку, что повышает ее возбудимость. Локальный ответ является экспериментальным феноменом, однако по перечисленным выше свойствам он близок к таким явлениям, как процесс местного нераспространяющегося возбуждения и возбуждающего постсинаптического потенциала (ВПСП), который возникает под влиянием деполяризующего действия возбуждающих медиаторов.
Потенциал действия
Потенциал действия (ПД) возникает на мембранах возбудимых клеток под влиянием раздражителя пороговой или сверхпороговой величины, который увеличивает проницаемость мембраны для ионов натрия. Ионы натрия начинают входить внутрь клетки, что приводит к уменьшению величины мембранного потенциала – деполяризации мембраны. При уменьшении МП до критического уровня деполяризации открываются потенциалозависимые каналы для натрия и проницаемость мембраны для этих ионов увеличивается в 500 раз (превышая проницаемость для ионов калия в 20 раз). В результате проникновения ионов натрия в цитоплазму и их взаимодействия с анионами разность потенциалов на мембране исчезает, а затем происходит перезарядка клеточной мембраны (инверсия заряда, овершут) – внутренняя поверхность мембраны заряжается положительно по отношению к наружной (на 30 – 50 мВ), после чего закрываются натриевые каналы и открываются потенциалозависимые калиевые каналы. В результате выхода калия из клетки начинается процесс восстановления исходного уровня мембранного потенциала покоя – реполяризация мембраны. Если такое повышение проводимости для калия предотвратить введением тетраэтиламмония, который избирательно блокирует калиевые каналы, мембрана реполяризуется гораздо медленнее. Натриевые каналы можно блокировать тетродотоксином и разблокировать последующим введением фермента проназы, который расщепляет белки.
Таким образом, в основе возбуждения (генерации ПД) лежит повышение проводимости мембраны для натрия, вызываемое ее деполяризацией до порогового (критического) уровня.
В потенциале действия различают следующие фазы:
1. Предспайк – процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
2. Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
3. Отрицательный следовой потенциал – от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
4. Положительный следовой потенциал – увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
Изменения возбудимости при возбуждении