В качестве отправной точки для реконструкции Либермана и Крелина послужили представления о существовании корреляции между определёнными деталями морфологии базикраниума (основания черепа), с одной стороны, и положением голосовых органов (в том числе гортани и языка), с другой. Имевшиеся в то время сравнительно-анатомические данные свидетельствовали, в частности, о том, что при слабо изогнутом основании гортань расположена высоко, а при сильно изогнутом низко. Считалось, что последняя черта характерна лишь для людей, причём справедливо отмечалось, что у младенцев гортань расположена так же высоко, как у многих животных (что даёт им и животным возможность есть и дышать практически одновременно), и лишь в 2–3 года начинает опускаться (что позволяет лучше и разнообразнее артикулировать звуки, но создаёт риск подавиться). У шапелльского неандертальца конфигурация основания черепа в некоторых отношениях напоминала таковую у современных младенцев, что во многом и предопределило выводы, к которым пришли Либерман и Крелин в своей знаменитой работе.
Впервые серьёзные причины усомниться в полученных ими результатах появились после того, как в конце 80-х годов французским антропологом Ж.-Л. Эймом была выполнена новая реконструкция черепа из Ля Шапелль-о-Сен. Она отличалась от старой реконструкции М. Буля, которой пользовались Либерман и Крелин, и одно из отличий как раз состояло в большей степени изгиба основания черепа. У Эйма получалось, что по этому признаку шапеллец вполне вписывался в рамки вариабельности, наблюдаемой у современных людей[182].
Однако хотя очередной пересмотр анатомии многострадального неандертальца из Ля Шапелль (предыдущий, о котором говорилось в главе 1, был предпринят в 50-е годы, но касался в основном посткраниального скелета) заставил усомниться в правильности реконструкции Либермана и Крелина, он никак не решал проблему с черепами других его собратьев, которые упрямо не желали менять малый изгиб основания на большой. По всем измерениям получалось, что в целом неандертальцы в этом отношении стоят в стороне от «магистральной линии» эволюции гоминид. Конечно, по крайней мере у некоторых из них изгиб базикраниума был сильней, чем у австралопитеков, мало отличавшихся по этому признаку от человекообразных обезьян, и даже сильней, чем у ранних гомо эректус, имевших зачаточный изгиб, но зато явно слабей, чем у гомо гейдельбергенсис (Петралона, Брокен Хилл). У последних фиксируется уже полный изгиб, близкий к наблюдаемому на черепах современных людей, а вот у неандертальцев базикраниальный угол заметно больше (т. е. основание черепа менее изогнуто), чем у гомо сапиенс (рис. 7.4)[183]. Что бы это могло означать? Неужели регресс?
Результаты проведённого недавно изучения полутора сотен черепов обезьян и гоминид, представлявших 29 разных видов включая неандертальцев и гомо сапиенс, помогают ответить на этот вопрос. Целью исследования было проверить гипотезу, согласно которой степень изгиба базикраниума у приматов в значительной мере зависит от размеров мозга и лица и от соотношения этих размеров. Проведённые измерения и вычисления подтвердили исходное предположение. Оказалось, что чем больше мозг, тем, при прочих равных условиях, больше изгиб основания, но если росту мозга сопутствует увеличение размеров лица, то это ведёт к некоторому уменьшению изгиба. Такой характер взаимодействия трёх названных величин хорошо объясняет, почему при примерно равном объёме мозговой полости изгиб базикраниума у неандертальцев меньше, чем у современных людей[184]. Ведь лицо у них, как мы помним из главы 2, было намного больше, чем у нас.
Ну а как же в этом случае быть с корреляцией между степенью изгиба основания черепа и положением гортани? Что же получается: большое лицо = малый изгиб базикраниума = высоко расположенная гортань = проблемы с членораздельной речью? Не слишком ли сложная конструкция? Согласен, слишком. Но в ней нет необходимости. К сожалению, исходные посылки, на которых строились реконструкции Либермана и многие более поздние исследования речевых способностей ископаемых гоминид, не выдержали проверки временем. Во-первых, оказалось, что в процессе вокализации у многих животных гортань может опускаться так же низко, как она постоянно опущена у людей, а значит, её высокое «стационарное» положение само по себе не является препятствием для речи. Во-вторых, как теперь известно, у некоторых млекопитающих и стационарная позиция гортани не отличается от человеческой. У разных видов оленей, у коал, а также у всех кошек рода Panthera (т. е. у львов, тигров, леопардов и ягуаров) она, как и у нас, опущена постоянно, а не только во время рёва или рычания. Низкое положение гортани обеспечивает понижение тона издаваемых звуков, что создаёт у получателей сигнала преувеличенное представление о размерах и силе обладателя «баса», и, возможно, именно в этом и заключается основной адаптивный смысл данного признака (вспомним ломающиеся голоса подростков). Наконец, в-третьих, как это уже ясно из «во-первых» и «во-вторых», корреляция между степенью изгиба основания черепа и положением гортани оказалась совсем не обязательной[185]. Это было дополнительно подтверждено экспериментами (на крысах), а также наблюдениями над детьми: у них, когда гортань опускается, изменений конфигурации базикраниума не происходит. Таким образом, получается, что будь даже изгиб основания черепа у неандертальцев таким же, как у шимпанзе, т. е. почти никаким, это всё равно не означало бы отсутствия у них речи. Впрочем, и наличия таковой это тоже, разумеется, не означало бы. Вообще же главный вывод из сказанного совсем не радостен: мы лишились основного из тех немногих скелетных индикаторов способности к членораздельной артикуляции звуков, на которые, как казалось, можно было смело опираться в попытках реконструировать речевой потенциал ископаемых гоминид.
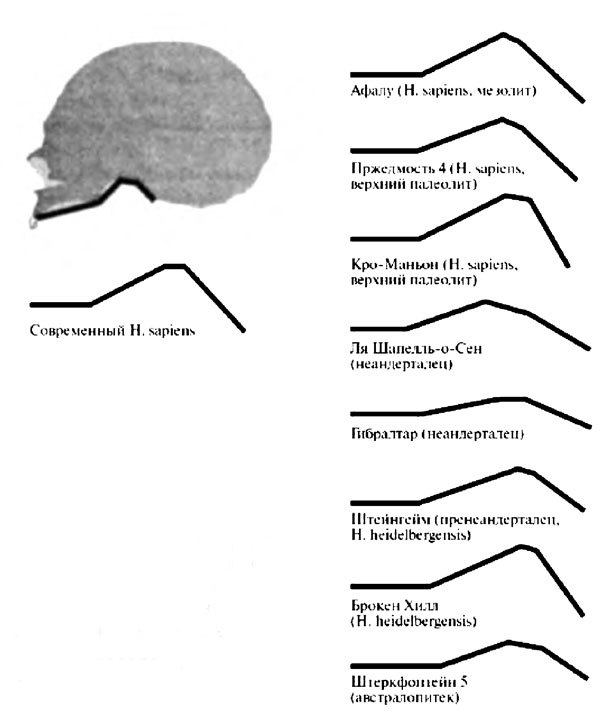
Рис. 7.4. Изгиб основания черепа у современного человека (слева) и ископаемых гоминид (справа). Как видно, неандертальцы явно отклоняются от общей тенденции (источник: Laitman et al. 1978)
К счастью, не все открытия имеют столь печальные последствия. Иногда они кое-что проясняют, ничего при этом не руша. Одно такое открытие, имеющее самое непосредственное отношение к вопросу о фонетическом «репертуаре» неандертальцев, было сделано в пещере Кебара в Израиле в 1983 г. Речь идёт о находке подъязычной кости (гиоида) одного из представителей этого вида. Его останки, погребённые около 60 тыс. лет назад, отличались от большинства современных им костяков очень хорошей сохранностью. Череп, правда, отсутствовал (объяснение чему даётся в конце следующей главы), но зато остальной скелет был представлен почти полностью, причём все его части располагались в правильном анатомическом порядке. Сама подъязычная кость, занимающая при жизни человека позицию в мягких тканях над гортанью и связанная с мускулами горла и языка, лежала, как ей и следует, рядом с нижней челюстью. Её изучение, предпринятое израильскими и французскими антропологами, показало, что по своему строению она ничуть не отличалась от аналогичной кости современных людей. Позже такое же заключение было сделано и относительно двух подъязычных костей пренеандертальцев из пещеры Сима де лос Уэсос[186], тогда как найденный недавно гиоид австралопитека афарского оказался точно таким же, как у шимпанзе[187]. Конечно, строго говоря, сама по себе идентичность формы ещё не означает, что подъязычная кость и гортань занимали у людей из Симы и Кебары такое же низкое положение в горле, как у нас, но результаты осуществлённой недавно виртуальной реконструкции позволяют предполагать, что так оно и было. Эта реконструкция проводилась методом трёхмерной геометрической морфометрии по опорным точкам на черепах и нижних челюстях людей и шимпанзе, для которых были получены сканы головы и шеи. В итоге авторы работы (опубликованы пока лишь тезисы) пришли к выводу, что у неандертальцев гиоид, скорее всего, занимал по отношению к черепу, шее и позвоночному столбу позицию близкую к той, что он занимает у современных людей, и что, таким образом, этот вид обладал одной из предпосылок, необходимых для человеческой речи[188].