Таким образом, палеогенетические данные, как и анатомические, говорят в пользу обособленного видового статуса неандертальцев. Оценки времени происхождения этого вида (или, точнее говоря, времени, когда жил последний общий предок всех «классических» неандертальцев), полученные методом молекулярных часов, укладываются в интервал от 150 до 250 тыс. лет назад, а с учётом стандартного отклонения — от 120 до 350 тыс. лет назад. Разделение линий неандертальцев и современных людей произошло, судя по первым результатам применения того же метода, скорее всего, где-то в интервале от 500 до 700 тыс. лет назад, или, если брать крайние даты, от 320 до 850 тыс. лет назад[80]. Самые последние расчёты времени этого события, основанные на данных по полностью расшифрованной мтДНК из Виндии, дали дату 660±140 тыс. лет[81]. Она хорошо согласуется с палеоантропологическими данными и является, вероятно, наиболее реалистичной. Вместе с тем, следует иметь в виду, что результаты датировок, получаемых методом молекулярных часов, зависят от целого ряда плохо поддающихся проверке допущений. Таковы, в частности, допущения относительно скорости накопления различий (мутаций)[82], постоянства этой скорости, «нейтральности» мутаций, т. е. отсутствии влияния отбора на их накопление, размера популяций и их демографической истории. Есть и другие факторы, осложняющие расчёты и заставляющие воспринимать молекулярные датировки с осторожностью. Они, безусловно, полезны, и во многих случаях остаются единственным способом оценить время интересующих нас филогенетических событий, но точность их не стоит переоценивать.
Литература
Эволюционная история гоминид: Вишняцкий 2004, 2005б; Зубов 2004; Фоули 1990; Arsuaga and Martínez 2006; Cameron and Groves 2004; Cela-Conde and Ayala 2007; Finlayson 2009; Johanson and Edgar 1996; Klein 1989; Lewin 2005; Lewin and Foley 2004; Tattersall and Schwartz 2000; Wood 2005; Wood and Lonegran 2008.
Происхождение и эволюция неандертальцев: Дробышевский 2004, 2006; Зубов 1999, 2004: 245–340; Хрисанфова 1997; Arsuaga et al. 1996, 1997; Condemi 2000; Deanetal. 1998; Harvati 2007; Hawks and Wolpoff 2001; Howell 1951; Hublin 1998, 2007, 2009; Rightmire 2008; Rosas et al. 2006; Schrenk and Müller 2009: 51–71; Tattersall 1995: 130–147; Tattersall and Schwartz 2006; Trinkaus 1988.
Таксономический статус неандертальцев: Харитонов 1973; Harvati 2003b; Harvati et al. 2004; Schillaci and Froehlich 2001; Smith 2010; Tattersall 2007; Wolpoff 2009.
Генетика неандертальцев: Овчинников и др. 2009; Briggs et al. 2009; Goodwin and Ovchinnikov 2006; Green et al. 2006, 2008, 2010; Hebsgaard et al. 2007; Lalueza-Fox et al. 2006, 2007; Noonan et al. 2006; Orlando et al. 2006; Pennisi 2009; Serre and Pääbo 2006.
Глава 4
На краю света
Почему неандертальцы стали такими, какими они стали? Что привело к появлению у них тех анатомических особенностей, которые отличают их от гомо сапиенс и других гоминид и позволяют считать особым биологическим видом? Вопрос этот очень труден, но не неразрешим. Антропологам есть что на него ответить. Ответ, конечно, будет далеко не исчерпывающим, но, как ни странно, гораздо более полным, чем в том случае, если бы мы поставили тот же вопрос применительно к своему собственному виду. Хотя очень многие специфические черты неандертальской морфологии пока ещё не получили сколько-нибудь удовлетворительного объяснения, причины и общий функциональный смысл эволюции неандертальцев понятны сегодня всё же лучше, чем причины и смысл большинства тех анатомических изменений, которые сопутствовали появлению гомо сапиенс. В целом, конечно, мало кто сомневается в том, что направление биологической эволюции обоих видов, особенности их анатомии и образа жизни в значительной мере зависели от природных условий, в которых им приходилось жить и к которым они должны были приспосабливаться. Однако если характер задач, которые окружающая среда могла ставить перед неандертальцами, более или менее ясен, то о гомо сапиенс этого пока не скажешь. Ведь становление первых происходило на севере, в краю с довольно суровой, переменчивой и очень требовательной природой, а вторые формировались в тропиках, где климат неизмеримо мягче, а перепады его далеко не столь резки и непредсказуемы, как в высоких широтах. К разговору о гомо сапиенс нам ещё предстоит вернуться в одной из следующих глав, а сейчас попытаемся представить, каковы были естественные условия существования неандертальцев, и как конкретно сказалось влияние этих условий на их анатомии и внешнем облике.
Мир вокруг них
Для начала несколько слов о погоде. О том, какой она была во времена неандертальцев и как удалось об этом узнать. Откуда, в частности, стало известно, что климат Европы в те далёкие времена был суров и переменчив? И можно ли эту — слишком уж общую — характеристику как-то развернуть, наполнить подробностями, конкретными деталями?
Для реконструкции природной обстановки минувших эпох используются самые разные методы и материалы. О климате и ландшафтах плейстоцена очень многое можно узнать, изучая ископаемые зёрна пыльцы растений, кости животных, формы рельефа земной поверхности, характер чередования и структуру геологических напластований, химический состав ископаемых почв и т. д. Например, по пыльце, сохраняющейся в погребённом состоянии на протяжении сотен тысяч лет, можно определить, каков был характер и состав растительности в том или ином регионе в тот или иной период. Эти данные, в свою очередь, могут много сказать о климате, поскольку каждый вид трав, кустарников и деревьев требует определённой температуры и влажности. У животных также всегда были и есть свои климатические и ландшафтные предпочтения, и знание их не менее полезно для палеоклиматических реконструкций, чем знание «привычек» растений. Находка, скажем, костей песца и северного оленя в древнем слое где-нибудь в верховьях Дуная или в среднем течении Дона — это явное свидетельство того, что здесь некогда было намного холоднее, чем сейчас, и что зона тундры заходила в соответствующий период далеко на юг.
Особенно большое значение для палеогеографических реконструкций приобрёл в последние десятилетия метод, основанный на анализе изотопного состава раковин некоторых морских микроорганизмов (фораминифер), хорошо сохраняющихся в ископаемом состоянии. С помощью глубоководного бурения получают колонки донных отложений, накапливавшихся сотни тысяч лет, а затем сравнивают соотношение концентрации стабильных (т. е. нерадиоактивных) изотопов кислорода 18O и 16O у фораминифер из разных слоёв. Поскольку первый из этих изотопов тяжелее второго, и содержащие его молекулы воды испаряются медленнее, то его роль в построении раковин (или доля в составе раковин) была неодинакова в холодные и тёплые эпохи. Во время оледенений, когда огромные массы испарившейся влаги не возвращались обратно в мировой океан, а оказывались «запертыми» в ледниках, концентрация 18O в морской воде увеличивалась, тогда как в периоды межледниковий она, наоборот, уменьшалась. Таким образом, по изменению соотношения 18O/16O можно проследить общий ход колебаний температуры на протяжении длительных периодов времени. Выделенные таким образом эпохи истории климата называют кислородно-изотопными стадиями или морскими изотопными стадиями (рис. 4.1).
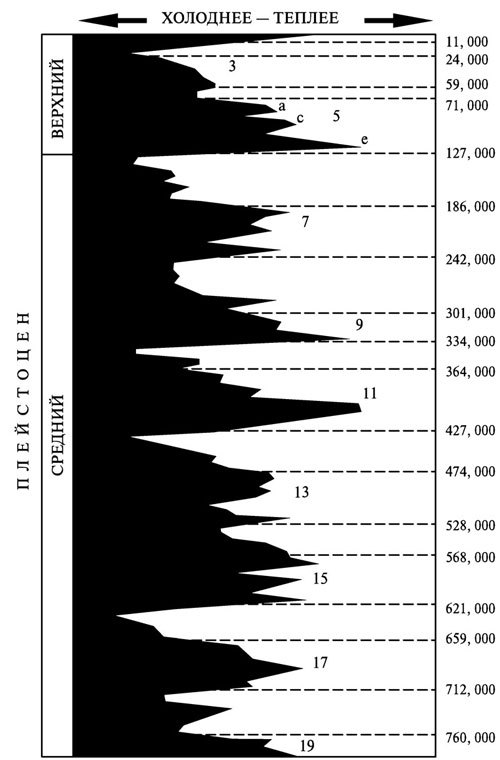
Рис. 4.1. Общая направленность колебаний климата в среднем и верхнем плейстоцене, т. е. в период после проникновения людей в Европу. Цифрами от 1 до 19 пронумерованы кислородно-изотопные стадии (чётные номера соответствуют похолоданиям, нечётные — потеплениям), показаны также их хронологические рамки
80
Green et al. 2006; Noonan et al. 2006.
81
Green et al. 2008.
82
Как уже говорилось, для расчёта темпа накопления мутаций необходимы надёжные и хорошо датированные палеонтологические реперы (точки отсчёта), которые на самом деле имеются лишь в редких случаях. Например, для людей в качестве такого репера часто используют, как уже говорилось, время расхождения линий гоминид и шимпанзе, но датировка этого события зависит от того, кого мы принимаем за первого гоминида. Если сахельантропа, то расчёт нужно вести от даты 7 млн. лет назад, а если оррорина, то 6 млн. лет назад, а ведь есть ещё и иные варианты. Авторы первых генетических датировок происхождения Homo sapiens (или, точнее, времени существования последнего общего «митохондриальоного предка» ныне живущих людей) в качестве репера использовали время дивергенции американоидов и монголоидов, которую тогда (четверть века назад) относили в прошлое на 12 тыс. лет (Cann et al. 1987). Сегодня почти никто не сомневается, что это событие (т. е. заселение Америки) произошло, по крайней мере, на 2 тыс. лет раньше, а многие готовы увеличить его древность и ещё больше.