* Аминопептидаза М, аминопешчдаза N, аланинаминопептидаза.
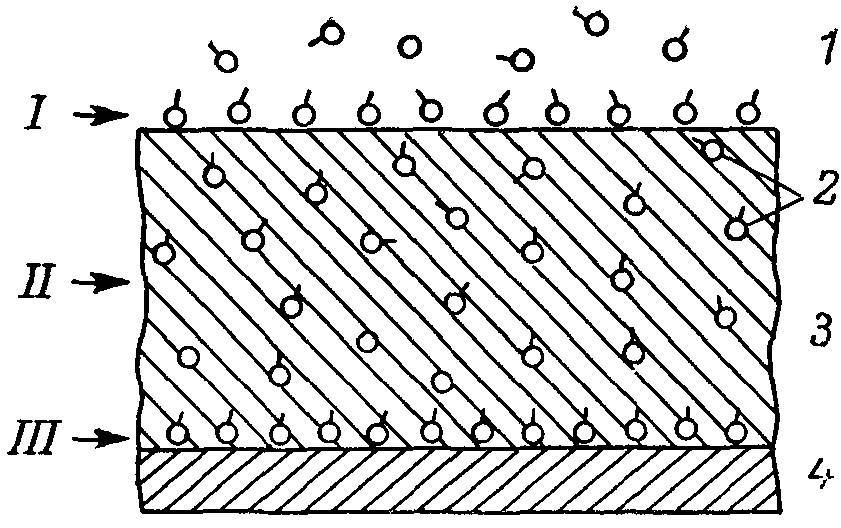
Рис. 5. Упрощенная схема распределения адсорбированных ферментов на поверхности гликокаликса (I), в гликокаликсном пространстве (II) и на липопротеиновой мембране (III) кишечной клетки.
1 — полость тонкой кишки; 2 — ферменты;3 — гликокаликс; 4 — мембрана.
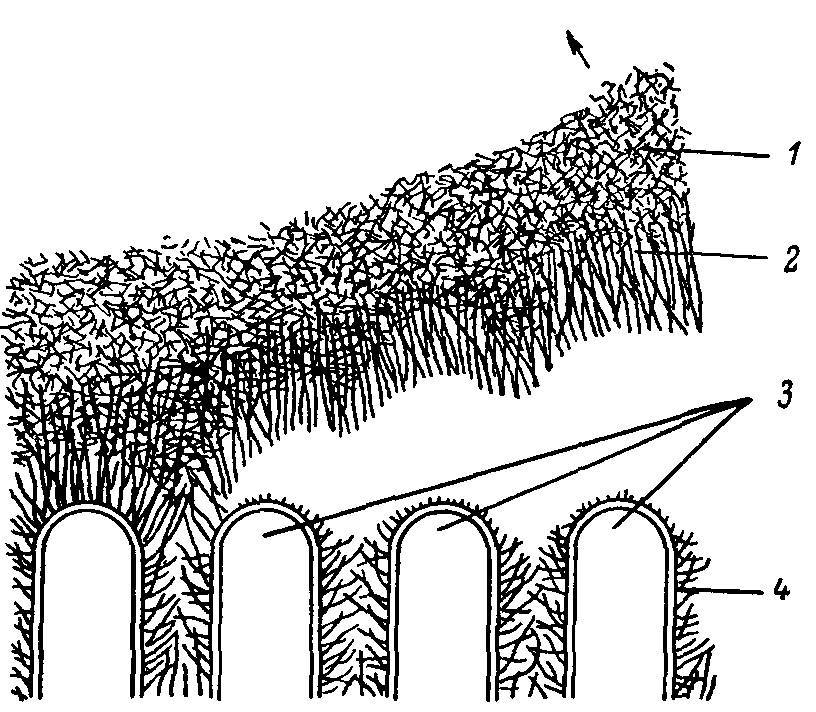
Рис. 6. Схема отделения апикального гликокаликса от липопротеииовой мембраны кишечной клетки.
1 — агаровая реплика; 2 — апикальный гликокаликс; 3 — микровор-синки; 4 — латеральный гликокаликс.
Рис. 7. Щеточная кайма кишечной клетки крысы.
А— продольный разрез апикальной зоны интактной клетки; виден гликокаликс на внешней (апикальной) и боковой (латеральной) поверхностях микроворсинок. В — продольный разрез апикальной зоны клетки после отделения агаровой реплики; гликокаликс на внешней поверхности микроворсинок отсутствует, видны неповрежденная липопротеиновая мембрана клетки и латеральный гликокаликс. 80 000х.
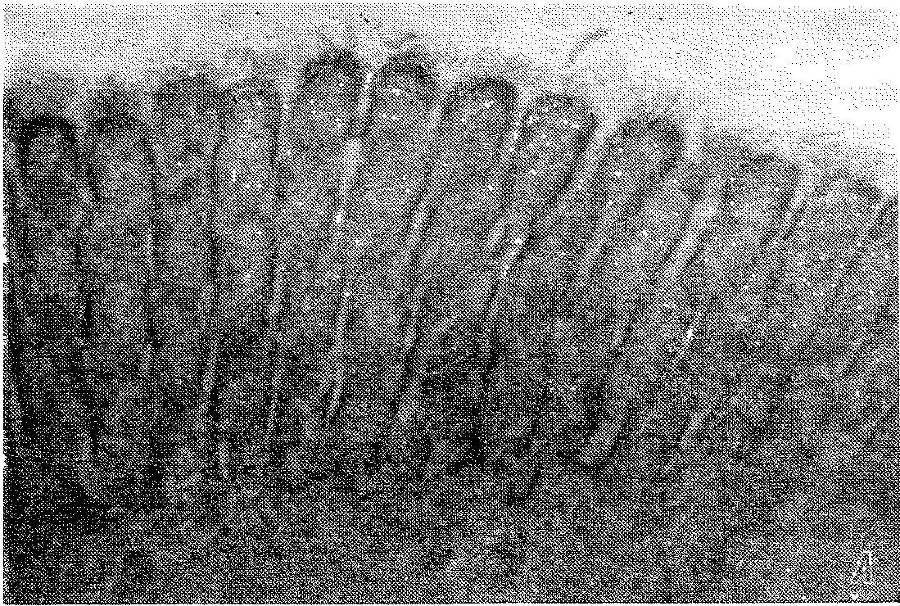
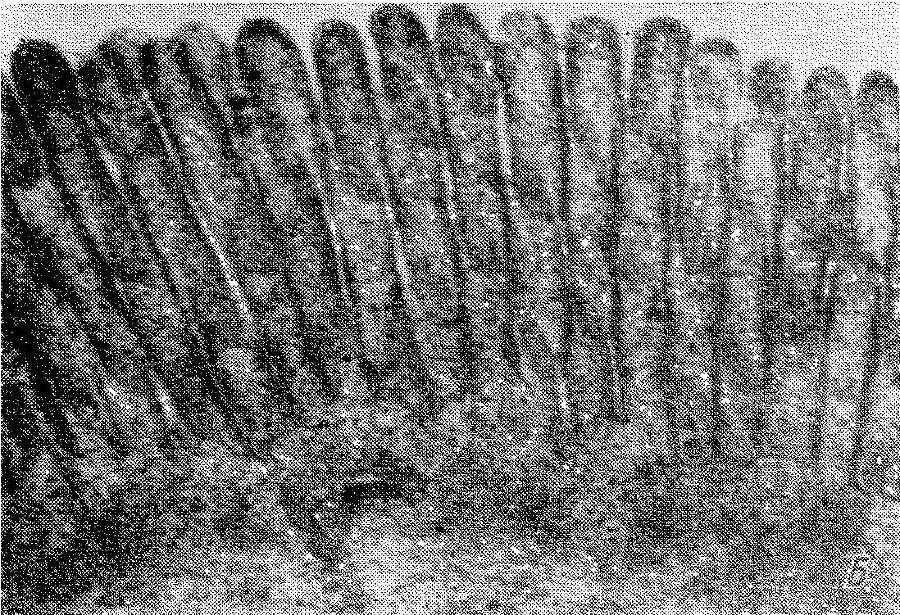
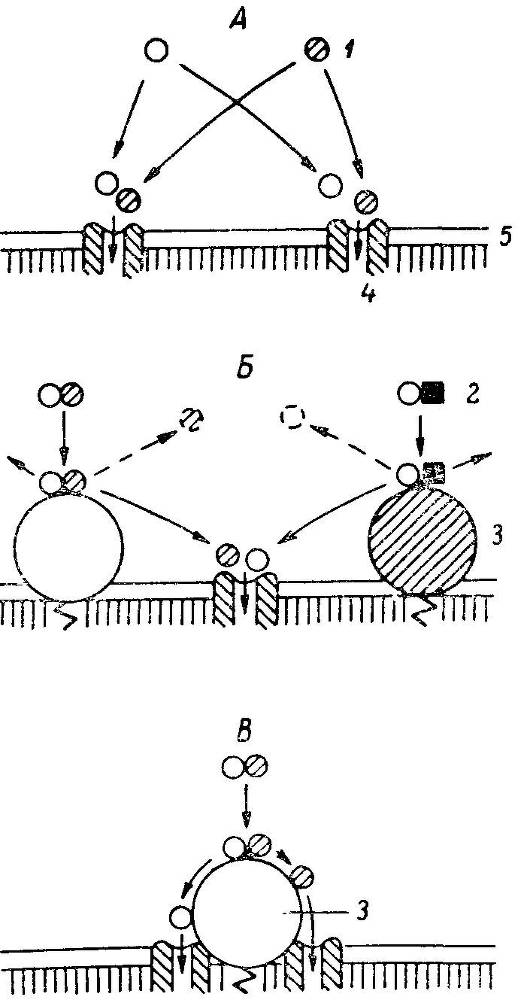
Рис. 8. Роль ферментно-транспортных комплексов в предотвращении конкуренции между мономерами на стадии всасывания.
А — конкуренция между мономерами за общий вход в транспортную систему; Б — конкуренция между финальными продуктами гидролиза за общий вход в транспортную систему; В — ферментно-транспортный комплекс: передача конечных продуктов гидролиза с фермента на вход в транспортную систему (без конкуренции). 1 — мономер; 2 — димер; 3 — фермент; 4 — транспортная система; 5 — мембрана.
Несколько лет назад нами впервые препаративно отделен апикальный гликокаликс от плазматической мембраны кишечных клеток крыс без повреждения этой мембраны (рис. 6, 7). Было обнаружено, что в апикальном гликокаликсе, отделяющем мембрану от внеклеточной среды, сосредоточено около 60% панкреатической амилазы, более 80% трипсина и около 20% химотрипсина, адсорбированных на структурах кишечной слизистой оболочки. Следовательно, примерно 40% амилазы, 20% трипсина и 80% химотрипсина, адсорбированных на этой оболочке, могут быть локализованы в латеральном гликокаликсе, т.е. в межмикрозорсинчатом пространстве, а также, возможно, частично на липопротеиновой мембране. В этих же экспериментах продемонстрировано, что такие собственно кишечные ферменты, как сахараза, гамма-амилаза, ди- и трипептидазы, связаны преимущественно с липопротеиновой мембраной. Однако щелочная фосфатаза, рассматриваемая как трансмембранный интегральный фермент, присутствует в довольно значительных количествах (до 20%) в апикальном гликокаликсе. (Сходные данные получены па курах). Кроме того, выявлено, что в апикальном гликокаликсе содержится 3.6% лейцинариламидазы, менее 2% пролилглициндипептидазы и менее 1% глицилпролинди-пептидазы.
Благодаря локализации кишечных ферментов на липопротеиновой мембране в непосредственной близости от транспортных систем мембранное пищеварение обеспечивает сопряжение конечных этапов переваривания и начальных этапов всасывания. Это достигается в результате специальной организации пищеварительных и транспортных функций клеточной мембраны в виде своеобразного пищеварительно-транспортного конвейера, способствующего передаче конечных продуктов гидролиза с фермента на вход в транспортную систему и предотвращению конкуренции между ними за обладание входом в последнюю (рис. 8). Иными словами, мембранный фермент и транспортная система образуют олигомерный комплекс, между частями которого существуют кооперативные и аллостерические взаимодействия (рис. 9, 10).
Мембранное пищеварение наблюдается у организмов на всех уровнях эволюционного развития, т.е. является универсальным механизмом. Оно обнаружено у всех млекопитающих, включая человека, у птиц, рыб, амфибий, круглоротых, а также у беспозвоночных животных, в том числе у насекомых, ракообразных, моллюсков, у различных паразитирующих форм. Существуют данные о наличии мембранного пищеварения у дрожжей, бактерий и в корнях растений.
Для правильной оценки мембранного пищеварения существенны следующие факторы.
1. Ферменты, реализующие этот процесс, относятся к структурированным. В связи с этим возможна пространственная организация как ферментных, так и транспортных систем, объединяющих заключительные этапы переваривания и начальные этапы всасывания.
2. Структурирование ферментов приводит к изменению их свойств. Так, отделение ферментов от мембраны меняет их каталитические и регуляторные характеристики.
3. Зона мембранного пищеварения обладает особыми физико-химическими свойствами, в частности такими как pH, концентрация органических и неорганических ионов, неперемешиваемый слой жидкости.
4. Мембранное пищеварение осуществляется в стерильной зоне, недоступной для бактерий, что предотвращает поглощение последними легко усвояемых низкомолекулярных пищевых веществ.
5. Благодаря ферментному аппарату гликокаликсный слой превращается в высокоспецифический фильтр. Через этот слой проникают те вещества, для которых на поверхности и внутри гликокаликсного пространства имеются адекватные ферменты, но не проникают другие вещества с такими же размерами молекул.
6. Для понимания эффективности мембранного пищеварения и трансмембранной проницаемости важны сведения о сократительной функции щеточной каймы, регулируемой кальцием. Показано, что микрофиламенты микроворсинок, выполняющие эти функции, содержат актин и миозин и связаны с апикальной мембраной кишечных клеток.
7. Следует учитывать не только процессы синтеза кишечных ферментов и их включения в состав апикальной мембраны кишечных клеток, по и скорость их деградации. (Более подробно см. гл. 5).
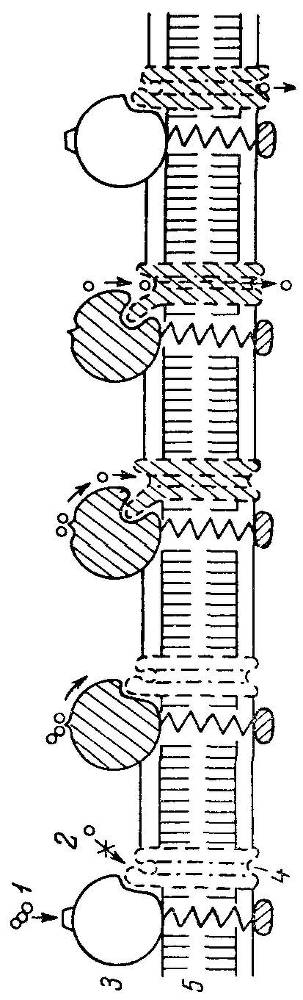
Рис 9. Схема последовательных конформационных взаимодействий и транспортной частей комплекса.
1 - субстрат; 2 - продукт; 3 - трансмембранный фермент; транспортная система; 5 - мембрана
2.4. Схема переваривания пищи как сочетание трех основных типов пищеварения
После обнаружения мембранного пищеварения классическая схема ассимиляции пищи претерпела существенные изменения. Согласно классическим представлениям, пищевые вещества — нутриенты, способные к всасыванию и ассимиляции, освобождаются в результате ферментативного гидролиза сложных органических соединений за счет внеклеточного (полостного) и внутриклеточного типов пищеварения. При этом усвоение пищевых веществ происходит в два этапа: полостное пищеварение—всасывание. Согласно современной схеме, усвоение пищи реализуется не в два, а в три этапа: полостное пищеварение—мембранное пищеварение—всасывание с более или менее выраженным компонентом внутриклеточного пищеварения. Таким образом, мембранное пищеварение, занимая по функциональной позиции промежуточное положение между полостным пищеварением и всасыванием, является акцепторным механизмом по отношению к полостному гидролизу и донорным по отношению к всасыванию. Следовательно, полостное пищеварение без мембранного не имеет существенного значения, так как всасывание без предварительного мембранного гидролиза невозможно из-за отсутствия адекватных субстратов. Вместе с тем нельзя недооценивать роль полостного пищеварения, так как обычные пищевые продукты не способны проникать в зону щеточной каймы и гликокаликсное пространство без обработки в пищеварительных полостях.