Насколько можно судить, экскреторный аппарат насекомых может обеспечить удаление избытка воды и солей и строгую экономию воды. Это достигается вариациями в распределении насосов и каналов и интенсификацией поглощения солей и воды (рис. 39).
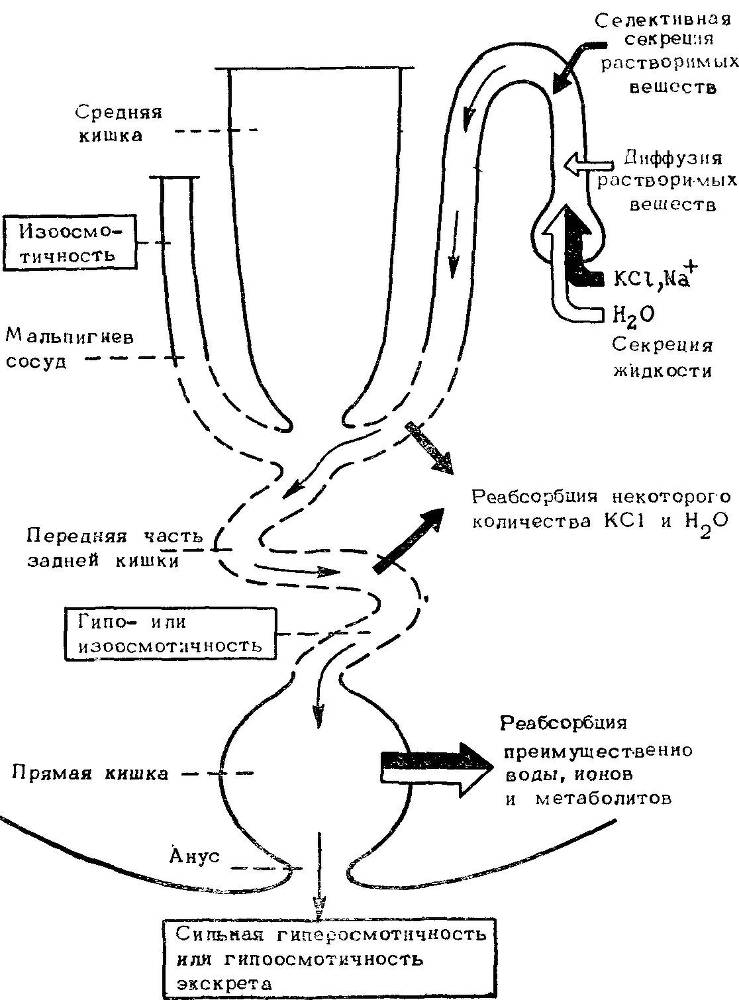
Рис. 39. Схема типичной экскреторной системы насекомых.
Указаны основные транспортные процессы и конечная осмолярность жидкости, покидающей каждый сегмент. Тонкие стрелки — выделение мочи, толстые — активный транспорт, светлые — пассивный транспорт.
Таким образом, адаптация выделительной системы насекомых и млекопитающих основана на использовании идентичных или близких функциональных блоков. Последние, однако, принадлежат к структурам, происходящим из различных зародышевых листков и, казалось бы, с различной эволюционной специализацией. Еще более замечательным является то, что многие регуляторные функции млекопитающих и насекомых осуществляются с помощью идентичных молекул.
Одним из примеров перемещения гормонально активных веществ и изменений контура эндокринной регуляции может служить формирование двух источников АКТГ — нейроэндокринного (гипоталамо-гипофизарная система) и желудочно-кишечного. Ясно, что в ходе эволюции подобные взаимоотношения могли складываться лишь как результат перераспределения функциональных блоков.
Интересно, что одни и те же пептиды могут синтезироваться в тканях, имеющих различное фило- и онтогенетическое происхождение. Так, холецистокинин и гастрин могут синтезироваться в эпителиальных клетках экто- и энтодермального происхождения и в нервных клетках.
АТФ — основной макроэрг в механизме первичной энергизации, в ходе эволюции может выступать также в роли нейротрансмиттера. При этом АТФ включен в синаптические везикулы, и его эффект, как и других нейротрансмиттеров, реализуется с помощью экзоцитоза. Не менее выразительным примером является использование для передачи сигналов синапсов аминокислот, которые накапливаются в гранулах и действуют при экзоцитозе. В настоящее время известны глициновая и глутаминовая сигнализации, а также рецепторы для связывания аминокислот.
Перестановка функциональных блоков в пределах клетки, их перераспределение между различными частями одного органа или различных органов не связаны с какими-либо ограничениями. Действительно, увеличение или уменьшение количества рецепторов инсулина, изменение типов секреции и количества выделяемого гормона не нарушают взаимоотношений рецептора и лиганда, но создают ряд новых ситуаций в таких взаимодействиях. Некоторые изменения могут оказаться полезными. Так, инсулинпродуцирующие клетки, первоначально локализованные в тонкой кишке (например, у круглоротых), затем переместились в поджелудочную железу. Глюкагонпродуцирующие клетки, также первоначально локализованные в тонкой кишке у высших позвоночных, частично сохранились в этой области. В то же время другая часть таких клеток транслоцировалась в поджелудочную железу.
Н. Трак в 1973 г. высказал гипотезу о существовании универсального кишечного прогормона. Автор предположил, что кишечные гормоны произошли в результате мутации из проинсулинподобной молекулы-предшественника в ходе эволюционного развития. При дупликации гена из проинсулинподобной молекулы возникли две молекулы проинсулина, одна из которых стала предшественником инсулина, а другая (при дальнейшей мутации и дупликации гена) — гастрина и секретина. Секретинподобная молекула стала, в свою очередь, предшественником секретина и глюкагона.
Еще более ярким примером того, как концепция функциональных блоков дает ключ к пониманию ранее таинственных явлений, может быть происхождение различных ядов. Этот пример демонстрирует перемещение функциональных блоков в филогенезе.
6.3.3. Происхождение ядов. Гормоны и яды
Еще Ч. Дарвин обращал внимание на трудности в объяснении происхождения ядов. Недавно мною высказано предположение, что объяснение происхождению ядов следует искать в концепции универсальных функциональных блоков. Допускается, что яды появляются в результате постепенного формирования новых физиологически активных факторов ради защиты, нападения и добывания пищи и образуются в специализированных органах благодаря транслокации гена или его экспрессии из одного оперона в другой. При этом первоначальная физиологическая роль данного фактора могла заключаться в поддержании регуляторных функций организма. Тот же фактор, продуцируемый в больших количествах и вводимый при защите и нападении, вызывает токсикоз у жертвы. Например, фосфолипазы — физиологические ферменты в составе мембран, при введении в кровь с ядом змей провоцируют ее гемолиз и другие деградационные процессы, приводящие к смерти. Точно так же функцию физиологических регуляторов в своем организме могут выполнять батрахотоксин и другие вещества. В то же время накопленные в слюнных железах и введенные в организм человека, они вызывают токсический эффект.
При анализе молекулярной структуры показано, что яды, как и гормоны, представляют собой пептиды и в ряде случаев имеют сходную структуру. В коже амфибий обнаружено много активных пептидов с широким спектром действия. Кожные яды амфибий присутствуют в качестве гормонов в желудочно-кишечном тракте млекопитающих, а также в качестве рилизинг-факторов в их организме. Примером этому может служить бомбезин — один из наиболее известных ядов кожи лягушки. Этот пептид у млекопитающих, в том числе у человека, по отношению к гормонам желудочно-кишечного тракта служит рилизинг-фактором, т.е. гормоном, обеспечивающим освобождение из клеток других гормонов. Точно так же бомбезин и некоторые другие гормоны и кожные яды присутствуют в центральной нервной системе высших организмов.
Таким образом, яды кожи лягушки — вещества, сходные с уже известными гормонами, а во многих случаях идентичные им. Подобное заключение сделано в отношении других ядов. В частности, недавно обнаружены два пептида. Один из них — саувагин, изолированный из кожи лягушки, с физиологическими эффектами на сердечно-сосудистую систему и переднюю долю гипофиза теплокровных, другой — дерморфин — вещество с мощными опиатными эффектами, в том числе аналгезийными.
Важно, что все обнаруженные кожные пептиды амфибий имеют функциональные аналоги с гормонами энтеронейрональной оси млекопитающих. Так, функциональные эффекты церулеица сходны с таковыми холецистокинина и гастрина, эффекты тахикинина — с субстанцией Р и т.д. В кишке и мозгу выявлена бомбезинподобная иммунореактивность (табл. 17). Еще более поразительно, что структура пептидов кожи амфибий близка, а в некоторых фрагментах идентична структуре гормонов человека и высших животных. Подобные совпадения не случайны и отражают единство происхождения пептидов. Это означает, что гипотеза происхождения ядов в результате генетических экспрессий сформировавшегося физиологически активного вещества в других органах получает подтверждения.
Семейство пептидов Кожи лягушки в аналогов пептидов в кишке в мозгу млекопитающих
| Кожа | Кишка | Мозг |
| Тахикинины | Субстанция Р | Субстанция Р |
| (эледозин, физалемин) | Физалеминподобные пептиды | Физалеминподобные пептиды |
| Церулеин | Xолецистокинин/гастрины | Xолецистокинин-8 |
| Брадикинины | ? | Брадикининподоб-ная иммунореактивность |
| Боыбезины | Бомбезинподобные пептиды | Бомбезинподобные пептиды |
| Ксенопсин | Нейротензин | Нейротензпн |
| Ангиотензин | Ангиотензины | Ангиотензины |
| Гормон, освобождающий тиреотропин | Гормон, освобождающий тиреотропин | Гормон, освобождающий тиреотропин |
| ВИП | ВИП | ВИП |
| Саувагин | ? | ? |
| Дерморфин | Опиоидные пептиды | Опиоидные пептиды |