Внутриклеточные сигналы. На уровне сигнальных рецепторных блоков концепция их универсальности и специализации на основе рекомбинации находит новое подтверждение. Например, секреторные процессы в ацинарных клетках поджелудочной железы контролируются системой нервных и гормональных сигналов. В частности, выброс секреторных гранул происходит под влиянием холецистокинина и ацетилхолина. Холецистокинин длительное время рассматривался как специфический стимулятор секреции ферментов. Однако позднее было обнаружено, что этот механизм не специфичен, а включен в систему вторичных мессенжеров — цАМФ и Са2+. Так, холецистокинин контролирует многие цАМФ- и Са2+-зависимые процессы, в том числе внутренней секреции, стимулируя выделение ряда гормонов, и действует аналогично медиатору. Многие гормоны выполняют также функции нейротрансмиттеров. Эту роль могут играть гастрин и холе-цистокинин; серотонинподобные вещества, вызывающие возбуждение в межнейронных синапсах; АТФ — скорее всего в тормозящих нейронах; субстанция Р, выполняющая функции возбуждающего, а энкефалин и соматостатин — тормозящего нейротрансмиттеров. Предполагается также нейротрансмиттерная функция ВИПа (рис. 29).
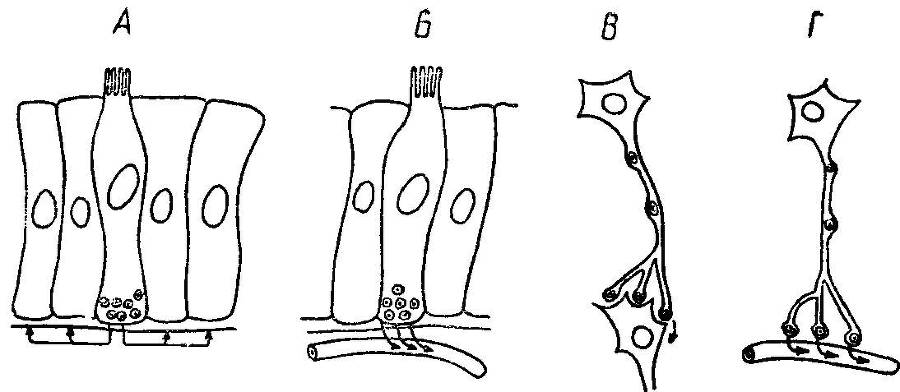
Рис. 29. Схема способа передачи пептидов, действующих в качестве паракринных мессенжеров (А), кишечных гормонов ( Б ), нейротрансмиттеров (В) и нейрогормонов (Г).
Те же самые молекулы могли бы функционировать в каждой из систем одного и того же организма.
Принцип работы рецепторных блоков можно проиллюстрировать на примере рецепторно-аденилатциклазного комплекса. Такой комплекс осуществляет передачу сигналов путем их ретрансляции с помощью аденилатциклазы. Сущность процесса сводится к образованию комплекса рецептор—гормон, что приводит к стимуляции активности аденилатциклазы, локализованной на внутренней стороне мембраны. Активация аденилатциклазы первичным мессенжером связана с его взаимодействием с рецептором, каталитической и, возможно, промежуточной субъединицами фермента. При стимуляции активности фермента происходит увеличение образования цАМФ, что вызывает цепную реакцию с отрицательной обратной связью, приводящую к ее выключению. Ниже приведен список гормонов, стимулирующих или подавляющих активность аденилатциклазы (табл. 11).
Таблица 11
Гормоны, оказывающие влияние на аденилатциклазную активность различных тканей
| Гормон | Ткань-мишень |
| Катехоламины | Многие ткани |
| Глюкагон | Печень, жировая ткань, В-клетки поджелудочной железы |
| АКТГ (адренокортикотропный гормон) | Кора надпочечников, жировая ткань |
| ТСГ (гормон, стимулирующий выделение тиреотропина) | Щитовидная железа |
| Паратиреоидный гормон | Почки, костная ткань |
Ясно, что с помощью различных соотношений стандартных рецепторных и циклазных блоков может быть достигнута высокая избирательность регуляции различных органов (рис. 30). Вместе с тем при некоторых условиях даже небольшое количество определенных рецепторов может быть важным при развитии адаптационных (например, адаптационно-компенсаторных) реакций или при развитии патологических процессов, например вследствие побочных эффектов гормональных веществ.
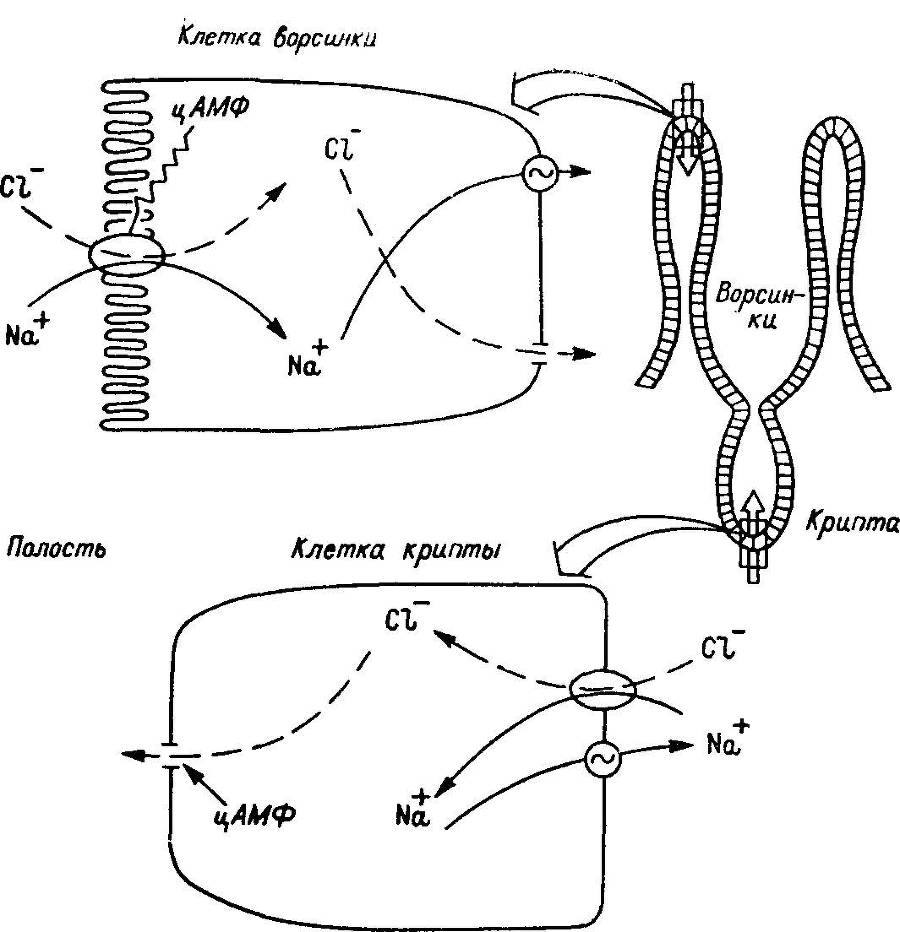
Рис. 30. Предполагаемая схема действия цАМФ на транспорт ионов в ворсинках и криптах тонкой кишки млекопитающих.
Показаны два секреторно-чувстительных процесса, разделенных пространственно: абсорбтивный электронезависимый, локализованный в клетках ворсинок, и электрогенный секреторный, локализованный в клетках крипт. Эта же схема иллюстрирует предполагаемую цАМФ-медиироваввую секрецию электролитов в тощей кишке. Антиабсорбтивные (в клетках ворсинок) и секреторные (в клетках крипт) эффекты ввутриклеточных мессенжеров разделены. Гипотеза может иметь отношение ко всей секреции, стимулируемой циклическими нуклеотидами и Са2+, так как цАМФ и Са2+ -ионофоры оказывают эффект на транспорт, сходный с эффектами цАМФ.
Таким образом, наряду с воспринимающим блоком — собственно рецептором — существует эффекторный блок — аденилатциклаза. Эта система универсальна и присутствует в клетках всех органов и тканей. Система циклических нуклеотидов имеется не только у позвоночных, большинства беспозвоночных, включая первичноротых, но и у одноклеточных организмов. У последних, так же как и у грибов, цАМФ выполняет роль передатчика сигналов не внутрь клеток, а от одной особи к другой. Следовательно, цАМФ первично возникает на весьма ранних стадиях формирования эукариотов как типичный гормон и, сохраняясь как химический мессенжер и постепенно подвергаясь интернализации, превращается во вторичный мессенжер.
Таблица 12
Внутриклеточные ферменты, регулируемые кальмодулином
| Фермент | Локализация фермента |
| Фосфодиэстераза циклических нуклеотидов | Мозг, щитовидная железа, легкие, эмбрионы цыпленка, околоушная железа, гепатома, околощитовидные железы |
| Аденилатциклаза | Мозг, поджелудочная железа, сперма, прокариоты |
| Гуанилатциклаза | Тетрахимена |
| цГМФ-протеинкиназа | Мозг |
| Киназа легких цепей миозина | Гладкие мышцы, скелетная мускулатура, сердечная мышца, почки, кровяные тельца |
| Са2+, Mg2+-ATФaзa | Эритроциты и адипоциты (плазматическая мембрана), мозг (синаптическая плазматическая мембрана), сердечная мышца (саркоплазматический ретикулум) |
| Динеин | Реснички тетрахимены, растения (мембраны микро-сом) |
| Фосфорилаза-кина за | Скелетная мускулатура, кровяные тельца, сердечная мышца |
| Синтаза-киназа гликогена | Печень |
| НАД-киназа | Растения, морской еж |
| Фосфопротеин-фосфатаза | Печень, мозг |
| Фосфолипаза А2 | Кровяные тельца |
| Триптофан-5-монооксигеназа | Мозг |
| Сукцинатдегидрогеназа | Печень |
| О-Метилтрансфераза | Слизевик |
| N-Метилтрансфераза | Мозг |
Одновременно с цАМФ существует и взаимодействует с ним другой регуляторный механизм — система Са2+—кальмодулин. Многие Са2+-зависимые процессы обусловлены присутствием внутриклеточного белка — кальмодулина. Последний служит основным Са2+-связывающим белком как мышечных, так и немышечных клеток. Многие известные процессы, опосредованные Са2+, включают кальмодулин как промежуточный комплекс. Тот факт, что кальмодулин служит внутриклеточным рецептором Са2+, позволяет думать, что Са2+ играет роль вторичного мессенжера подобно цАМФ. Они имеют много общего в механизме действия. С влиянием кальмодулина на метаболизм циклических нуклеотидов и гликогена, а также на транспорт Са2+ связана регуляция многих ферментов в клетках различных типов (табл. 12). Это относится также к контролю сократительной активности клеток. Кальмодулин является компонентом цитоскелета кишечных клеток и обнаружен в микроворсинках этих клеток и клеток плаценты, в клетках мозга, матки, семенников, скелетной мускулатуры и т.д. Кальмодулин и Са2+ универсальны и характерны для всех эукариотов и прокариотов. У эукариотов кальмодулин связан в единый регуляторный блок с системой циклических нуклеотидов. Сходный феномен наблюдается и у простейших, для которых характерна развитая система Са2+—кальмодулин, взаимодействующая с аденилатциклазной системой.