Гликокаликс определяет ряд других важнейших функций клеток: «узнавание», связь молекул определенных типов (т.е. специализированные акцепторные и рецепторные функции), иммунологическое дифференцирование, межклеточные взаимодействия и т.д. По-видимому, он играет роль в связывании поверхностного слоя слизи. Гликокаликс является не только диффузионным барьером, препятствующим проникновению бактерий, но и специфическим барьером, определяющим проницаемость пищевых веществ. Благодаря ему апикальная мембрана практически недоступна для высокомолекулярных соединений типа ксенобиотиков. В то нее время гликокаликс обеспечивает высокую проницаемость для молекул пищевых веществ, так как в его пространстве адсорбированы пищеварительные ферменты панкреатического происхождения.
От состояния гликокаликса и содержащихся в нем ферментов могут зависеть многие нарушения функций желудочно-кишечного тракта. В частности, нарушение адсорбции панкреатических ферментов имеет значение в формировании явлений малнутриции, а атрофия гликокаликса может явиться причиной повреждающего действия токсических агентов химуса на липопротеиновую мембрану.
Быстрое обновление гликокаликса обеспечивает эффективное функционирование щеточной каймы как пористого реактора, так как благодаря сбрасыванию «зрелого» гликокаликса создается своеобразный эффект постоянной очистки пор. Недавно показано активное сбрасывание гликокаликса в результате активации фибриллярных структур микроворсинок терминальной сети.
Следует подчеркнуть, что гликокаликс представляет собой универсальную структуру и встречается у клеток всех организмов от амебы до человека, включая эритроциты и нейроны. В то же время гликокаликс характеризуется вариабельностью свойств не только в клетках разного типа, но и в одной клетке. Например, он сильно развит на апикальной поверхности микроворсинок кишечных клеток, менее — на их латеральной поверхности и лишь биохимически (но не структурно) определяется на базальной мембране.
Актомиозиновый комплекс. Сократительные структуры характерны для возбудимых тканей (мышечной и нервной) и контролируют форму клеток. Однако оказалось, что эти структуры присутствуют в самых различных клетках. Недавно некоторые блоки сократительной системы описаны в невозбудимых клетках, и в том числе в кишечных.
На рис. 27 представлена сократительная система апикальной части кишечной клетки. Как можно видеть, сердцевина микроворсинки занята фибриллярными структурами, которые достигают апикальной области цитоплазмы, где сливаются с поперечной фибриллярной структурой — терминальной сетью. Последняя, согласно существующим представлениям, выполняет функцию опоры для щеточной каймы, принимает участие в регуляции подвижности мембраны, а также разделяет клетку на два компартмента — щеточнокаемный и цитозольный.
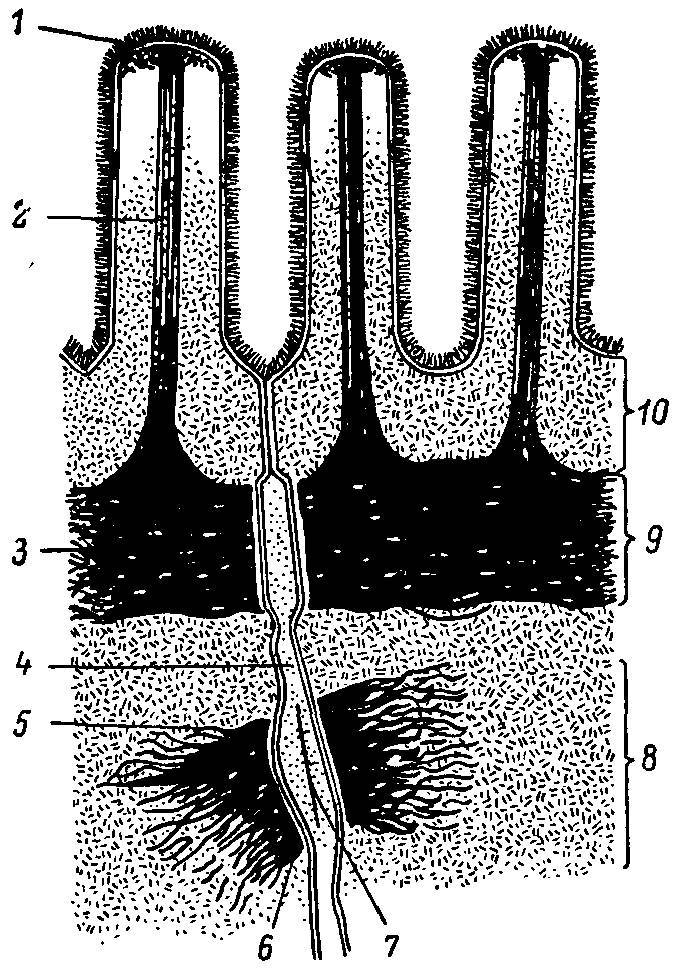
Рис. 27. Схема мембраны микроворсинок и апикальных соединительных комплексов латеральной мембраны кишечных клеток.
1 — гликокаликс; 2 — микрофиламенты; 3 — терминальная сеть; 4 — межклеточное пространство; 5 — тонофиламенты; в — плотное пятно; 7 — промежуточная плотная линия; 8 — десмосома; 9 — промежуточный контакт; 10 — плотный контакт.
Возникает вопрос о функциях сократительных структур в невозбудимых клетках. Нами еще в начале 60-х гг. было высказано предположение, получившее подтверждение, что благодаря актомиозиновому комплексу реализуются расслабления и сокращения микроворсинок, что может существенно влиять на скорость мембранного пищеварения и всасывания. Недавно выявлена еще одна функция сократительного аппарата кишечных клеток — контроль их десквамации, обеспечивающий гомеостатирование барьерных свойств эпителиального пласта.
Наличие сократительных белков в различных клетках немышечной ткани говорит об их универсальности.
При этом актины немышечных и мышечных клеток сходны по своей структуре. Вероятно, актомиозиновый комплекс, присутствующий во всех эукариотических клетках и участвующий в двигательных процессах, включен в функционирование как возбудимых, так и невозбудимых тканей. Актомиозиновые структуры обнаружены также у большинства растений, у высших и низших грибов. Наличие актомиозиновых белков, по-видимому, служит той эволюционной основой, благодаря которой периодически возникают уникальные способности двигательного аппарата. Примером этому могут служить насекомоядные растения. Возникновение у них двигательных функций не кажется теперь столь неожиданным, так как присутствие сократительных структур характерно для клеток всех типов.
В сократительном блоке принцип регуляции является общим и мало меняется на протяжении эволюции от примитивных грибов до человека. Во всех случаях сокращение связано с активирующим действием Ca2+ и АТФазной активностью миозина. Характерно варьирование компонентов актомиозинового комплекса, при котором один из них может выпадать. Как правило, в филогенезе наиболее стабилен актиновый компонент, тогда как миозиновый по своему составу варьирует в широких пределах, так же как и их соотношения.
Гидролитические ферменты апикальной мембраны кишечных клеток. Эти ферменты — один из основных типов функциональных блоков в системе, выполняющей пищеварительные и транспортные функции в организме (т.е. в тонкой кишке). Особое значение имеют сравнительные исследования клеток тонкой кишки и почки. Эти органы, выполняющие различные физиологические функции, обладают такими идентичными ферментами, как мальтаза, аминопептидазы, эндопептидаза, щелочная фосфатаза, трегалаза и др. (табл. 7). Недавно из щеточнокаемных мембран клеток почки и плаценты выделены такие одноименные ферменты, как аланинаминопептидаза, мальтаза, сахараза, щелочная фосфатаза и др. Многие из этих ферментов обнаружены в мембранах клеток печени и поджелудочной железы, в кровеносной системе и т.д. В некоторых случаях удается показать близость или даже идентичность таких ферментов. Так, свойства щелочной фосфатазы клеток печени, почки, тонкой кишки, плаценты и сыворотки крови обладают значительным сходством. Обнаружено также сходство в иммунохимических характеристиках аланинаминопептидазы клеток поджелудочной железы, почки и печени человека.
Ферменты мембраны микроворсинок клеток почки и тонкой кишки млекопитающих
| Тривиальное наименование фермента | Наименование по КФ и № | Локализация |
| Аминопептидаза | Аминопептидаза (микросомная), 3.4.11.2 | Присутствует в почке и кишке |
| Аминопептидаза А | Аспартатаминопептидаза, 3.4.11.7 | То же |
| Дипептидилпептпдаза IV | Дипептидил пептидаза, 3.4.14.— или 3.4.21.— | » |
| Эндопептидаза | Нейтральна я пептидаза, 3.4.24.11 | » |
| Глутамилтрансфераза | Глутамилтрансфераза, 3.2.2.2. | » |
| Сахараза-изомальтаза | Саха раза-a-D-гликогидролаза, олиго-1,6-глюкозидаза, 3.2.1.48, 3.2.1.10 | Присутствует только в кишке |
| Мальтаза (глюкоамилаза) | a-D-Глюкозидаза, 3.2.1.20 | Присутствует в кишке и почке некоторых видов |
| Трегалаза | а-a-Трегалаза, 3.2.1.28 | Присутствует в почке и кишке |
| Лактаза-гликозилкерамидаза | β-D-Галактозидаза, 3.2.1.23, 3.2.1.62 | Присутствует только в кишке |
| Щелочная фосфатаза | Щелочная фосфатаза, 3.1.3 1 | Присутствует в почке и кишке |
Одни и те же ферменты, например щеточноrаемные гидролазы, могут присутствовать не только в мембране микроворсинок щеточной каймы, но и в латеральных и базальных мембранах, мембранах различных внутриклеточных органелл и даже в цитоплазме. Если присутствие щеточнокаемных ферментов в базолатеральной мембране свидетельствует об одном из физиологических путей от места синтеза к области их действия, то другая локализация, возможно, является полностью или частично результатом побочных эффектов. Однако