Поскольку после окончательного формирования мозга деление нейронов прекращается, необходимости в этой функции центросомы здесь нет. В центросомах нейронов, расположенных вблизи ядра, часто вовсе нет центриолей. Микротрубочки тянутся от центросом к окрестности пресинаптических окончаний аксона, а также в другую сторону, к дендритам и, через сокращающиеся актиновые нити, к дендритным шипикам, часто образующим постсинаптические окончания синаптической щели. Эти шипики способны расти и вырождаться, что, по-видимому, является существенным элементом общей пластичности мозга, благодаря которой система взаимных соединений в мозге подвергается непрерывным тонким изменениям. Насколько мне известно, существуют убедительные экспериментальные свидетельства важной роли микротрубочек в управлении пластичностью мозга.
Упомянем еще об одном любопытном факте. В пресинаптических окончаниях аксонов содержатся некие ассоциированные с миктротрубочками вещества, «работа» которых связана с высвобождением нейромедиаторов, а молекулы весьма примечательны с геометрической точки зрения. Эти вещества — клатрины— строятся из белковых тримеров (так называемых клатриновых трискелионов), этаких полипептидных трехлучевых звезд. Объединяясь в молекулу клатрина, трискелионы образуют геометрически правильные структуры, идентичные по общему строению многоатомным молекулам углерода, называемым «фуллеренами» (а также «бакиболами», или «мячами Баки» [54]) из-за их внешнего сходства со знаменитыми геодезическими куполами, которые проектировал и возводил американский архитектор Бакминстер Фуллер {91} . Клатрины, впрочем, гораздо больше фуллереновых молекул, поскольку одному атому углерода в фуллерене соответствует в клатрине целый трискелион, состоящий из нескольких аминокислот. Те клатрины, что заняты в высвобождении нейромедиаторов в синапсах, имеют форму усеченного икосаэдра— всем нам знакомого многогранника, по образу и подобию которого делают современные футбольные мячи (см. рис. 7.11и 7.12).
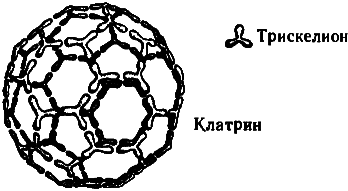
Рис. 7.11. Молекула клатрина (похожая общей структурой на фуллерен, но составленная не из атомов углерода, а из более сложных субструктур — белковых тримеров, называемых трискелионами). Изображенный на рисунке клатрин напоминает внешне обыкновенный футбольный мяч.
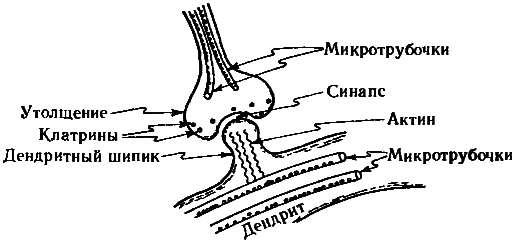
Рис. 7.12. Клатрины, подобные тому, что изображен на рис. 7.11, располагаются (вместе с окончаниями микротрубочек) в пресинаптическом утолщении аксона и, по всей видимости, участвуют в управлении интенсивностью синапса; также на интенсивность синапса влияют сокращающиеся актиновые нити в дендритных шипиках, управляемых микротрубочками.
В одном из предыдущих параграфов был поставлен важный вопрос: что управляет изменением интенсивности синапсов и определяет места размещения функционирующих синаптических связей? Учитывая имеющиеся свидетельства, можно уверенно предположить, что центральную роль в этих процессах играет цитоскелет. Как же это предположение может нам помочь в поиске невычислимой сущности разума? Пока что оно, похоже, говорит нам лишь о том, что потенциальная вычислительная мощность мозга оказывается гораздо большей, чем можно было бы ожидать, используй мозг в качестве простейших вычислительных блоков «цельные» нейроны.
В самом деле, если простейшими вычислительными блоками мы теперь будем считать димеры тубулина, то придется предположить, что потенциальная вычислительная мощность мозга просто неимоверно превосходит все то, что предполагали самые смелые теоретики от ИИ. Основываясь на «цельнонейронной» модели, Ханс Моравек в своей книге «Дети разума» [ 267] предположил, что человеческий мозг может в принципе достичь производительности порядка 10 14операций в секунду, но не более того; это при том, что в мозге имеется около 10 11 функционирующих нейронов, каждый из которых способен посылать примерно по 10 3сигналов в секунду (см. §1.2). Если же в качестве элементарного вычислительного блока взять димер тубулина, то следует учесть, что на каждый нейрон приходится около 10 7димеров; соответственно, элементарные операции теперь выполняются где-то в 10 6раз быстрее, в результате чего получаем 10 27операций в секунду. Возможно, производительность современных компьютеров и вправду уже начинает приближаться к первой цифре, 10 14операций в секунду (как весьма убежденно доказывают Моравек и его единомышленники), однако несмотря на все эти успехи, достичь в обозримом будущем производительности 10 27операций в секунду не представляется возможным.
Разумеется, можно смело утверждать, что мозг работает далеко не со стопроцентной «микротрубочковой» эффективностью, какую приведенные выше цифры предполагают. Тем не менее, ясно, что возможность «микротрубочкового вычисления» (см. [ 183]) позволяет совсем по-иному взглянуть на некоторые из аргументов в пользу неминуемого наступления эпохи искусственного интеллекта человеческого уровня. Можем ли мы теперь поверить хотя бы в то, что уже сегодня возможно {92} численно воспроизвести умственную деятельность червя нематоды, только потому, что мы вроде бы «закартографировали» и численно смоделировали его нервную систему? Как было отмечено в §1.15, умственные способности обычного муравья намного превосходят все то, что на настоящий момент реализовано посредством стандартных ИИ-процедур. Впору поинтересоваться, сколько же муравей выигрывает в производительности благодаря гигантскому массиву своих «микротрубочковых информационных нанопроцессоров», если сравнивать с тем, чего он смог бы добиться, располагай он лишь «переключателями цельнонейронного типа». Что до парамеции, то тут, как вы понимаете, оснований для предъявления иска нет.
Однако аргументы, представленные в первой части, предполагают гораздо более сильное заявление. Я утверждаю, что способность человека к пониманию выходит за рамки какой угодно вычислительной схемы. Если мозгом человека управляют микротрубочки, то в микротрубочковых процессах должно быть что-то принципиально отличное от простого вычисления. Я утверждал, что такая невычислимая активность должна быть следствием достаточно макроскопической квантовой когерентности, объединенной неким тонким образом с макроскопическим поведением — с тем, чтобы обеспечить возможность протекания в системе тех новых физических процессов, что придут на смену бытующей в современной физике паллиативной R-процедуре. В качестве первого шага мы должны выяснить, какова же подлинная роль квантовой когерентностив цитоскелетной активности.
7.5. Квантовая когерентность внутри микротрубочек
Есть ли у нас основания предполагать, что внутри микротрубочек существует квантовая когерентность? Вернемся ненадолго к обсуждавшимся в §7.1идеям Фрёлиха [ 131] о возможности феноменов квантовой когерентности в биологических системах. Он утверждал, что если энергия метаболической активности достаточно велика, а диэлектрические свойства задействованных в процессе материалов достаточно экстремальны, то существует возможность возникновения макроскопической квантовой когерентности, аналогичной той, что возникает в феноменах сверхпроводимости и сверхтекучести — иногда объединяемых общим термином конденсация Бозе—Эйнштейна— даже при относительно высоких температурах, какие, собственно, и характерны для биологических систем. Как выяснилось, не только метаболическая энергия достаточно велика, а диэлектрические свойства просто необыкновенно экстремальны (именно этот полученный в 1930-е годы поразительный экспериментальный результат и навел Фрёлиха на соответствующие размышления), но и имеется с некоторых пор даже прямое подтверждение предсказанных Фрёлихом внутриклеточных колебаний с частотой 10 11Гц [ 177].