Таким образом, появление белковой оболочки вокруг молекулы РНК явилось следующей ступенью эволюции материи от неживой формы к живой. Именно с появлением этой оболочки, можно говорить о качественно новом этапе эволюции материи — этапе эволюции живой материи. Стоячая волна мерности, возникающая во внутреннем объёме молекулы РНК или ДНК, являясь необходимым условием для возникновения жизни, не является достаточным в силу того, что, в первичном океане концентрация органических молекул была очень маленькой. И поэтому, без дополнительного накопления вблизи молекулы РНК органических молекул не могло быть и речи о постоянном захвате их этой молекулой с последующим распадом на материи, их образующие.
Возникает вопрос, каким образом у молекулы РНК могла появиться белковая оболочка? Какое чудо должно было произойти, чтобы это случилось? Ответ на эти вопросы, как ни странно, очень простой. Белки, как все органические молекулы, возникли в насыщенном растворе первичного океана, как следствие атмосферных электрических разрядов. Сами по себе белки являются большими молекулами, состоящиими порой из десятков тысяч атомов и при своём свободном (броуновском) движении в водах первичного океана, попадая в «поле тяготения» молекулы РНК, не могут быть втянуты во внутренний объём этой молекулы в силу того, что белки, сами по себе, представляют огромные молекулы и просто не могут поместиться во внутреннем объёме молекулы РНК. Но это не означает, что молекула РНК не влияет на молекулы белков. Создаваемый спиралью молекулы РНК радиальный перепад мерности, тем не менее, захватывает своим «тяготением» первичные структуры молекул белков и делает их своими постоянными «спутниками», как это происходит при захвате большим материальным телом (например, планетой) меньшего (Рис. 4.3.11).
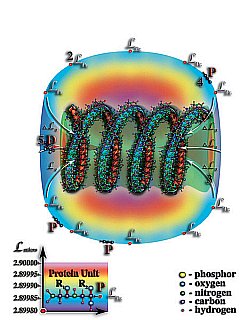
Рис. 4.3.11. Молекулы РНК или ДНК создают не только стоячую волну мерности во внутреннем объёме, но и создают вокруг себя перепад мерности микропространства. В результате этого, вокруг спиралей этих молекул образуются слои, имеющие тождественные уровни мерности. Влияние спиралей этих молекул на внешнее пространство значительно меньше влияния на мерность микропространства внутреннего объёма спиралей молекул РНК или ДНК. Тем не менее, на уровне микропространства спирали этих молекул выступают, как центры деформации микропространства. Молекулы ДНК и РНК на уровне микрокосмоса имеют двойственные свойства. Эти молекулы одновременно являются и аналогами «чёрных дыр» и звёздных систем на уровне микропростанства. Внутренний объём молекул РНК и ДНК проявляет свойства, аналогичные «чёрной дыре» макропространства, в то время, как внешний объём этих молекул проявляет свойства, аналогичные звезде. Все другие молекулы, попадая в поле притяжения этих «звёзд» — «чёрных дыр» микропространства или втягиваются во внутренний объём спиралей молекул РНК или ДНК, где распадаются на первичные материи их образующие, или оседают на уровнях тождественной мерности, которые возникают вокруг этих молекул. Первичные структуры молекулы белка, попадая в поле притяжения спиралей молекул РНК или ДНК, начинают оседать на уровне тождественной мерности LPr.
1. Физически плотная молекула ДНК или РНК.
2. Белковая оболочка.
3. Перепад мерности микропространства создаваемый внутренним объёмом молекулы ДНК или РНК.
4. Первичные структуры молекул белка.
P — аминокислоты белков.
R2 — свободные радикалы аминокислот белков.
LPr — уровень тождественной мерности первичной структуры молекулы белка.
И опять наблюдается удивительная параллель между макромиром и микромиром. Итак, захват и удержание молекулой РНК первичных структур молекул белков и стало тем условием, приведшим к формированию вокруг этой молекулы белковой оболочки. Со временем, число белков-спутников молекулы РНК возрастало, их «орбиты» были соизмеримы, в силу близких значений уровней собственной мерности. В результате чего, соседние белки-спутники оказывались друг от друга на таком расстоянии, что возникали условия для возникновения, так называемых, химических реакций между ними (Рис. 4.3.12).
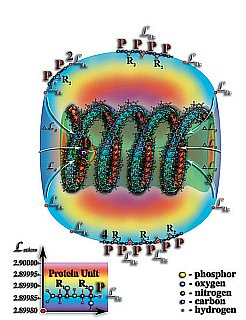
Рис. 4.3.12. Со временем, первичных структур молекул белка, захваченных полем притяжения молекул РНК и ДНК, становится всё больше и больше. Расположенные близко друг к другу, первичные структуры молекулы белка, посредством водородных связей и разнообразных связей между радикалами аминокислот, образующих первичные структуры белков, начинают создавать вторичную структуру белка.
В отличие от свободного синтеза белка из первичных стуктур белка, соединение последних происходит не произвольно. Удерживаемые полем притяжения спирали молекулы РНК или ДНК, первичные структуры белка вынужденно соединяются вдоль уровня тождественной мерности. В результате этого на уровне тождественной мерности LPr начинает формироваться белковая оболочка вокруг спирали молекулы ДНК или РНК. Уровень тождественной мерности вокруг спиралей молекул РНК и ДНК выступает, как организующее поле, заставляющее первичные структуры белка, захваченные «полем тяготения» спирали, соединяться в определённом порядке, как, например, силовые линии магнитного поля вынуждают крупицы металла располагаться по контурам этих силовых линий, которые, по сути своей, являются уровнями тождественной мерности, создаваемыми магнитом вокруг себя.
1. Физически плотная молекула ДНК или РНК.
2. Белковая оболочка.
3. Перепад мерности микропространства создаваемый внутренним объёмом молекулы ДНК или РНК.
4. Первичные структуры молекул белка.
P — аминокислоты белков.
R2 — свободные радикалы аминокислот белков.
LPr — уровень тождественной мерности первичной структуры молекулы белка.
Химические реакции между первичными структурами белков-спутников приводили к появлению устойчивых электронных связей между ними и они срастались в одно целое. Постепенно, вокруг молекулы РНК возникла сплошная белковая оболочка. Белки-спутники, в результате этого, заключили в свою собственную «тюрьму» своего захватчика — молекулу РНК (Рис. 4.3.13).
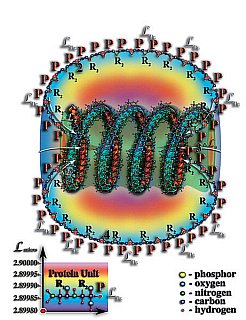
Рис. 4.3.13. Постепенно всё большее и большее число первичных структур белков захватывается «полем тяготения» спирали молекулы ДНК или РНК и вынужденно соединяются между собой на уровне тождественной мерности. Размер белкового слоя вокруг спирали молекулы ДНК или РНК постепенно растёт, и наступает момент, когда белковое поле полностью окружает спираль молекулы РНК или ДНК. Так появляется белковая оболочка у вирусов. Появление белковой оболочки вируса послужило началом новой эры эволюции материи — зарождению жизни. Белковая оболочка создала в своём внутреннем объёме условия значительно отличающиеся от условий вне её пределов. Она способствовала удержанию внутри себя органических и неорганических молекул, проникших через эту белковую сетку. Белковая оболочка вируса как бы процеживала воду первичного океана, собирая в своём внутреннем объёме растворённые в этой воде органические и неорганические молекулы. Подобная фильтрация морской воды позволяла накапливать органические молекулы в непосредственной близости от спирали молекулы ДНК или РНК. И, когда концентрация органических молекул достигала критического уровня, возникали условия для дублирования молекул ДНК или РНК и белковой оболочки. В результате этого процесса, возникала точная копия вируса. С этого момента можно говорить о зарождении жизни.
1. Физически плотная молекула ДНК или РНК.
2. Белковая оболочка.
3. Перепад мерности микропространства создаваемый внутренним объёмом молекулы ДНК или РНК.
4. Первичные структуры молекул белка.
P — аминокислоты белков.
R2 — свободные радикалы аминокислот белков.
LPr — уровень тождественной мерности первичной структуры молекулы белка.