Подобный процесс усиления боязни человека происходит почти всегда там, где человек проникает в ранее ненаселенную территорию и начинает охоту за животными местной популяции. Конечно, помимо отбора особей с наиболее резко выраженной пассивно-оборонительной реакцией и особей, которые легче других научились бояться человека, большую роль при указанной выше перестройке оборонительного поведения в популяции может играть традиция. Все многообразные сигналы тревоги, прямое подражание поведению родителей могут явиться наряду с отбором системой ненаследственной перестройки поведения популяции. Однако, если нет наследственно обусловленных предпосылок к проявлению пассивно-оборонительной реакции, один традиционный опыт, видимо, не в состоянии обусловить боязнь врага, истребляющего данную популяцию. Такой случай описан Ю. Гексли на примере гусей Фольклендских островов. Несмотря на интенсивное истребление человеком, они проявляли лишь небольшие признаки боязни человека, не предохраняющие популяцию от истребления. Наличие генотипических различий в формировании пассивно-оборонительной реакции является предпосылкой для перестройки оборонительного поведения популяции. Установлено, что усиление пассивно-оборонительной реакции происходит не за счет прямого наследственного закрепления реакций испуга уцелевших особей, а за счет естественного отбора наиболее трусливых особей или тех, которые имели генотип, способствующий наиболее быстрому формированию пугливости при их преследовании.
Без специального генотипического анализа трудно отдифференцировать наследственную и ненаследственную изменчивость поведения, традиции от поколения к поколению. Это издавна создавало условия для необоснованных допущений прямого наследования приобретенных навыков. Даже такой строгий и объективный исследователь, как И. П. Павлов, в очень осторожной форме допускал возможность наследования результатов индивидуального опыта. В 1913 г. он писал: «… можно принимать, что некоторые из условных вновь образованных рефлексов позднее наследственностью превращаются в безусловные» (С. 273). К тому же периоду относится его более определенное высказывание по этому вопросу в «Лекциях по физиологии»: «Передаются ли условные рефлексы по наследству? Точных доказательств этому нет, до этого наука еще не дошла. Но надо думать, что при длительном периоде развития прочно выработанные рефлексы могут становиться врожденными» (С. 85).[40] В начале 20-х годов И. П. Павлов поручил своему сотруднику Студенцову изучить наследование условных рефлексов у мышей. На эти опыты, не давшие положительных результатов, часто ссылались, причисляя И. П. Павлова к сторонникам наследования приобретенных признаков. Это вынудило И. П. Павлова в письме к Гуттену изложить свое отношение к этому вопросу (Правда. 1927. 13 мая). В течение всей дальнейшей жизни И. П. Павлов стоял на строго генетических позициях. Им была создана в Колтушах лаборатория по изучению генетики высшей нервной деятельности, перед зданием которой рядом с памятником Декарту и Сеченову был поставлен памятник Грегору Менделю. В качестве постоянного консультанта в своих генетических исследованиях И. П. Павлов пригласил крупнейшего генетика-невропатолога С. Н. Давиденкова, он консультировался также с Н. К. Кольцовым.
Генетическая работа велась путем селекции в отдельных семействах собак по типологическим особенностям их высшей нервной деятельности. Результаты этих исследований, показавшие роль генотипа в формировании типологических свойств высшей нервной деятельности, были опубликованы после смерти Павлова. Эти исследования показали, что генотипические факторы играют значительную роль в формировании типологических особенностей высшей нервной деятельности. В различных семействах собак, в которых производилась селекция в направлении степени силы (или слабости) процесса возбуждения, наблюдалась корреляция по этим особенностям нервной деятельности между братьями и сестрами в отдельных пометах собак: r=+34±0,1.
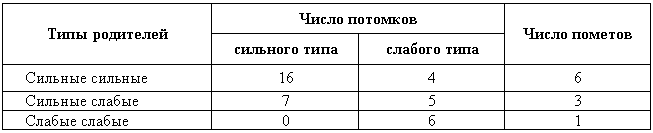
Результаты скрещиваний между собаками с сильной и слабой нервной системой, которые были начаты еще при жизни И. П. Павлова, сведены в табл. 2.
Степень силы нервной системы связана с полом: самцы (п = 31) обладают более сильной нервной системой, чем самки (п = 22). Вероятность соответствия Р(х2) оказалась меньше 0,05, что может быть признано статистически значимым. Вопрос о роли генотипических факторов в обучаемости исследуется начиная с известных работ Иеркс и Багга. Иеркс изучала обучаемость двух линий крыс: одну неинбридированную, другую инбредную из института Уистара. Результаты этой работы показали, что среднее время обучаемости неинбридированных линий несколько меньше, чем инбредных (52,25 урока у первых и 65,00 у вторых). Багг исследовал индивидуальные и семейные различия в поведении мышей при нахождении пути в довольно простом лабиринте. Сравнивалась линия белых мышей, которую Багг основал в 1913 г. (линия С, в настоящее время BALB/C), с линией желтых мышей. Оказалось, что среднее время обучаемости белых мышей за 15 опытов составляло 27,5±2,0 с с 9 ошибками за урок; время обучаемости желтых мышей равнялось 83,0±7,0 с с двумя ошибками за урок. Наблюдалось сходство в обучаемости особей одних и тех же пометов.
Детальное исследование роли генотипических факторов в выработке условных рефлексов было проведено Викари. В этом исследовании была изучена обучаемость японских танцующих мышей (Mus Wagneria asiatica) (инбридинг 20 лет), трех линий обычных мышей (Mus musculus), альбиносов Багга (инбридинг 14 лет), ослабленно-коричневых (инбридинг 17 лет) и ненормальноглазых (abnormal x-ray eyed) (инбридинг 6 лет). Оказалось, что каждая линия мышей имеет характерную для нее кривую обучаемости. Скрещивание между особями отдельных линий показало, что быстрая обучаемость доминирует над более медленной обучаемостью. Автор указывает, что характер расщепления во втором поколении позволяет предположить, что различие в обучаемости между коричневыми и альбиносами Багга обусловлено монофакторально, хотя нельзя отрицать и возможности более сложной картины наследования. Различия в обучаемости между линией Багга и японскими танцующими мышами обусловливаются, по мнению автора, наличием нескольких наследственных факторов. Проведенное исследование (выполненное на 900 мышах) указывает на большую роль генотипических факторов в быстроте обучаемости у мышей.
Однако найденные различия при анализе такого комплексного признака, каким является быстрота обучаемости, еще не доказывают генотипически обусловленных различий в интимных механизмах мозга, связанных с самим процессом обучения. Наличие генотипически обусловленных различий в безусловнорефлекторных реакциях изучаемых линий может в значительной мере определить найденные различия в обучаемости. В качестве подобного примера можно привести исключительно тщательную работу М. П. Садовниковой-Кольцовой. Проведя изучение обучения в лабиринте (хамптонкурт) у 840 крыс, автор путем селекции вывел две линии: одну быстро обучающуюся, другую медленно. Индекс быстро обучающихся крыс (логарифм времени, затраченного на 10 счетных опытов) равен 1,657±0,025, медленно обучающихся — 2,642±0,043. Разница между обоими индексами (Д=0,985±0,05) оказалась в 20 раз больше вероятной ошибки.
Дальнейший анализ показал, что найденные различия между обеими линиями крыс обусловливаются не различиями в их способности к выработке условных рефлексов, а большей пугливостью второй линии крыс (которая происходила в значительной мере от диких норвежских крыс). При обучении в аппарате Стона, в котором крыса подгонялась захлопывающимися дверцами и потому не могла из-за пугливости забиваться в угол лабиринта, обучение обеих линий шло одинаково.