Наблюдения на мышах показали, что ген, определяющий окраску шерсти, влияет и на поведение. Кроме того, описана связь одних поведенческих признаков с другими. Так, отбор мышей по «эмоциональности», которую оценивали частотой дефекации, привел к выведению двух линий, обозначаемых как реактивные и нереактивные. Оказалось, что в обеих линиях происходило однонаправленное изменение признака двигательной активности, по которому не велось отбора.
В некоторых случаях выявилась связь поведенческих признаков с морфологическими. Так, мыши линии C57BL, отличавшиеся высокой двигательной активностью, выраженностью ориентировочно-исследовательских реакций и малой эмоциональностью, были крупными, с густой шерстью на животе и коротким хвостом, а у мышей линии BALB/c все эти признаки имели противоположную направленность.
Фенотипические программы поведения гибридов отличаются от родительских и тем, что они оказываются более устойчивыми к внешним влияниям. Вместе с тем, когда эти программы складываются с участием влияния раннего опыта, они несут на себе отпечаток условий среды. Для мышей, например, особое значение имеют присутствие человека, шум, освещение, разнообразие окружения и действующих раздражителей, обозначаемые как обогащенная среда. Влияние условий развития мышей в обогащенной среде проявляется в большей степени на гибридах, чем на их родителях из инбредных линий (рис. 60).
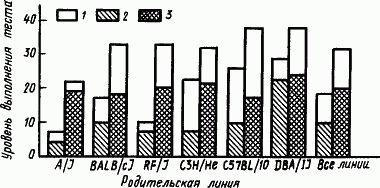
Рис. 60. Средние показатели успешного нахождения пищи мышами инбредных линий и гибридами от их скрещивания, выращенными в условиях стандартной и «обогащенной» среды (по Н. Гендерсону):
1 — эффект обогащенной среды, 2 — инбредная линия, 3 — гибрид
Исследование характеристик памяти на примере сохранения зрительного оборонительного условного рефлекса у крыс показало, что потомки наследуют свойства долговременной, но не краткосрочной памяти родителей.
Генетические программы пищевого поведения рогатого скота формировались в связи с условиями его содержания и кормления. Поэтому, например, у коров обнаруживаются характерные наследуемые породные особенности процесса жвачки. Различия программ пищевого поведения проявляются также в конкурентных отношениях с другими особями при подходе к кормушке. По этим показателям наблюдается прямая зависимость между такой активностью и размерами коров данной породы. Однако сравнительное исследование совместного поведения коров разных пород выявило, что наиболее активны в этом отношении коровы ангусской породы, им уступают шортгорны и на последнем месте оказались герефорды. Внутрипородные отношения перекрываются межпородными. Поэтому крупный герефорд уступает место у кормушки меньшему по размерам ангусу.
Значение наследственности в приручении диких животных наглядно показали наблюдения в питомнике серебристо-черных лисиц, распределение которых по классам прирученности (1 — высший, 2 — средний, 3 — низший) оказалось зависящим от числа ручных предков в их родословной (табл. 13).
Таблица 13. Распределение по классам прирученности, % (по Л.Н. Трут)

К наследуемым признакам относятся и некоторые патологические формы поведения. Так, отбор по чувствительности к звуковому раздражителю позволил вывести линию крыс, в которой 98–99 % особей реагировали на звук судорожными припадками. Характерной особенностью их поведения было двигательное возбуждение после прекращения звукового раздражения. Гибридологический анализ этой линии КМ (Л.В. Крушинский, Л.Н. Молодкина) показал, что такая наследуемая особенность поведения контролируется двумя генами.
Примером другой патологической формы поведения может служить подергивание лапок при выходе из эфирного наркоза у мух Drosophila melanogaster. Особи селекционированной по этому признаку линии были подвергнуты генетическому и нейрофизиологическому исследованию. Оказалось, что такой гиперкинез связан с геном, локализованным в X-хромосоме, и осуществляется нервными клетками грудного ганглия. Продолжение патологических разрядов нейрона даже при перерезке периферических нервов означает, что генерируемая им импульсация зависит от состояния его генетического аппарата.
Наблюдения за пищевым поведением инфузорий показали, что оно определяется их хеморецепцией, на которую оказывает регулирующее влияние аппарат ядра.
Дальнейшее более глубокое изучение механизмов генетического контроля поведения на уровне клетки проводили путем сопоставления мутантных признаков с состоянием физиологических, биохимических и электрофизических характеристик. Так, в опытах со скрещиванием мутантов инфузории Paramecium aurelia установлено, что изменение поведения парамеций путем перестройки движения ресничек при деполяризации или гиперполяризации мембраны контролируется генами определенной локализации. Генетический анализ с помощью спонтанных и индуцированных мутаций и испытания последних на аллельность и рекомбинации позволяют определить локализацию соответствующих генов и выяснить их связь с особенностями поведения.
Из поведения особей, связанного с половым отбором, складывается генотипический состав популяции. В результате миграции индивидов с различным набором генов и предпочтительного выбора самками гетерогенных самцов поддерживается генетический полиморфизм, открывающий широкие возможности возникновения новых признаков, которые закрепляются наследственностью. Таким образом, поведение и его изменчивость становятся одним из механизмов приспособительной эволюции вида.
Наследование типологических особенностей. История приручения диких предков домашних животных дает много поразительных примеров коренного изменения их темперамента. Например, такой контраст наблюдался между поведением коровы, спокойно пережевывающей жвачку, и ее предка — дикого тура, безудержного во взрывах дикой ярости. Подобные метаморфозы поведения достигались настойчивым отбором на протяжении многих веков. Животных отбирали с полезными для их использования типологическими особенностями. Скрещивание особей с наиболее выраженными полезными свойствами усиливало эти признаки. Так, были отобраны спокойные коровы, которые давали себя доить, смелые собаки, которые бросались на опасного хищного зверя, быстрые лошади, чутко реагирующие на малейшее движение всадника. Естественный отбор в природе приспосабливает темперамент животного к условиям его жизни.
Достигнутые в результате отбора признаки типа нервной системы передаются по наследству и проявляются в потомстве как врожденные. Это было показано в результате типологического изучения ряда поколений собак основного генетического стада в лаборатории экспериментальной генетики Института физиологии им. И.П. Павлова АН СССР.
Точные опыты определения типа нервной системы по стандарту, испытаний слюноотделительных рефлексов у родительских пар и потомства показали интересные закономерности наследования материнских и отцовских черт темперамента. Примером может служить следующее наблюдение.
У собаки-самки Пены наблюдали относительную слабость торможения. Она с большим трудом, после длительных применений, выработала дифференцировку, да и то неполную. Пену случили с кобелем Томбушем, имевшим отличную силу как возбудительного, так и тормозного процесса. О силе торможения у Томбуша можно было судить по тому, что ему достаточно было только трех применений дифференцировочного раздражителя, чтобы выработать абсолютную дифференцировку, которая оставалась полной все дальнейшее время работы. У Пены от Томбуша родилось три щенка: Прибой, Познай и Принц. Когда они подросли, типы их нервной системы стали изучать по стандарту типологических испытаний. Это исследование, продолжавшееся более двух лет, показало у всех потомков Пены и Томбуша унаследованную от матери четко выраженную относительную слабость торможения. Все они, как и Пена, с большим трудом вырабатывали дифференцировки, которые так и оставались нестойкими и неполными. Таким образом, типологические признаки одного из родителей оказались доминирующими, а признаки другого — рецессивными.