Анализ воздействия окружающей среды начинается с общеизвестной избирательной чувствительности рецепторов к раздражителям различной модальности. Такой качественный анализ, позволяющий воспринимать раздельно видимое, слышимое, пахнущее и т.п., дополняет количественный анализ, когда интенсивность воздействия кодируется амплитудой рецепторного потенциала, а затем частотой импульсов в афферентном нерве. Однако уже на этом первичном нервном уровне начинаются и процессы синтеза. Одним из них является интеграция сигналов от соседних рецепторов. Это впервые было показано на сложном глазе мечехвоста (Limulus) (Г. Хартлайн, Ф. Ратлифф, 1957). Разряд импульсов от одного омматидия тормозится при освещении другого, находящегося поблизости. Благодаря такому латеральному торможению освещение обоих рецепторов дает меньший эффект, чем каждого в отдельности. Оно выделяет контуры изображения и делает его более контрастным.
В сетчатке позвоночных обнаружены более сложные формы интеграции нейронных реакций, связанные со специфическим пространственным распределением их рецептивных полей. Типичными для такого распределения являются концентрические рецептивные поля (С. Каффлер, 1953). Для них характерны противоположные реакции центра и окружения. При этом выделяются так называемые on-нейроны, которые возбуждаются точечным засветом центра своего рецептивного поля, и off-нейроны, возбуждающиеся при выключении этого засвета; нейроны, которые возбуждаются и в том и в другом случае, обозначаются как on—off-нейроны. Засвет периферии рецептивного поля вызывает противоположные реакции, т.е. on-нейроны затормаживаются, a off-нейроны возбуждаются. Тормозное взаимодействие центра и периферии рецептивного поля осуществляется горизонтальными и амакриновыми клетками. В сигналах от нейронов дискретных концентрических рецептивных полей кодируются локальные перепады освещенности частей воспринимаемого изображения.
На более высоких уровнях обработки сенсорной информации происходит дальнейший синтез афферентных сигналов о признаках объекта. В связи с разной функцией нервных клеток они обнаруживают рецептивные поля разной конфигурации. На рис. 54, А показаны ответы нейрона зрительной коры кошки на точечный засвет в центре рецептивного поля и в зоне его периферии, а также на общее освещение. По наличию реакций в первом и во втором случаях этот нейрон относится к on — off-типу. Отсутствие реакций на общий засвет указывает на уравновешенность возбуждения в on- и торможения в off-нейронах. Примечательно, что рецептивное поле этого нейрона имело характерную вытянутую в виде полосы форму (рис. 54, Б). Значение такой формы рецептивного поля раскрывается в опытах с изменением наклона световой полоски, что приводит к уменьшению реакции при перекрытии полоской периферической части рецептивного поля.
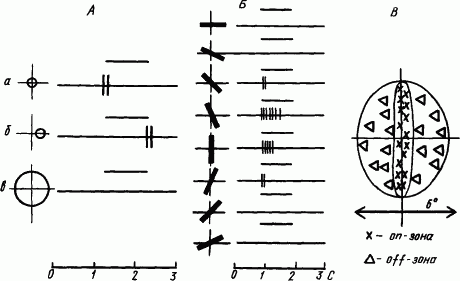
Рис. 54. Реакции нейронов зрительной коры мозга кошки на световое пятно (А, а, б), диффузное освещение (А, в) и световую полоску (Б) и общая форма вытянутого рецептивного поля (В):
слева от каждой нейрограммы указана форма стимула (кружок, полоска) и его локализация относительно рецептивного поля (крестик), над каждой нейрограммой чертой обозначена длительность стимуляции
Систематические исследования реакций нейронов зрительной коры мозга 17-го, 18-го и 19-го полей на световые стимулы разной формы и интенсивности, неподвижные и движущиеся, выявили важные закономерности формирования некоторых фрагментов нейрофизиологических механизмов зрительного восприятия (Д. Хьюбел, Т. Визел, 1974). Так, некоторые корковые нейроны наиболее активно реагировали на световую полоску, ориентированную вдоль вытянутого центра его рецептивного поля (рис. 54, Б). Как видно из рис. 54, наклоны полоски приводили к уменьшению ответа вплоть до его полного отсутствия. Такая избирательность коркового нейрона к ориентации линии находит объяснение в том, что на нем конвергируют сигналы от нескольких нейронов предыдущего этажа обработки зрительной информации (латерального коленчатого тела), и их концентрические рецептивные поля расположены в ряд, покрывающий световую полоску. Чем больше наклон полоски, тем меньше число вовлекаемых в реакции концентрических полей, а следовательно, и степень активации коркового нейрона. Если учесть, что в коре имеется множество нейронов с такими свойствами, но избирательно чувствительных к разной ориентации, то синтез их сигналов может дать описание всех видов линейных элементов контура воспринимаемого изображения. Такие нейроны называют простыми.
Другие нейроны зрительной коры имеют более сложную конфигурацию рецептивных полей, особенность которых состоит в отсутствии тормозного взаимодействия между участками поля. Поэтому они реагируют возбуждением на световой стимул, попадающий на разные его участки. Однако эти нейроны также строго избирательны к ориентации перепада освещенности. Вместе с тем их способность реагировать на стимул, последовательно проходящий ряд участков рецептивного поля, позволяет более точно оценивать его движение. При этом многие из них обнаруживают чувствительность лишь к одному направлению движения. Такие нейроны называют сложными.
Третий вид корковых нейронов, принимающих участие в обработке зрительной информации, характерен для рецептивных полей с комплексным возбудительным центром и тормозными зонами, ограничивающими размеры светового стимула, в частности длины его полоски. Эти клетки избирательно реагируют на ориентацию перепадов освещенности, линий и углов (главное, на их угловые размеры), оценивают направление движения стимулов. Такие нейроны называют сверхсложными.
Предполагают, что простые, сложные и сверхсложные клетки представляют три уровня синтеза результатов анализа зрительной информации. Свойства простой корковой клетки определяет конвергенция на ней сигналов от клеток латерального коленчатого тела, свойства сложной клетки — конвергенция на ней от простых клеток. Вопрос о происхождении сверхсложных клеток пока является дискуссионным.
Аналитико-синтетические процессы обеспечивают поэтапную обработку зрительной информации, направленную на восприятие целостного образа объекта. Изучение нейрофизиологических механизмов, осуществляющих эту обработку на разных этапах зрительного пути привело к представлению о четырех основных уровнях формирования целостного зрительного образа (В.Д. Глезер, 1985).
На первом уровне нейроны сетчатки и латерального коленчатого тела оценивают общую освещенность, выделяют сигналы из шума, подчеркивают контуры и дробят сетчаточное изображение на фрагменты.
На втором уровне нейроны 17-го поля коры определяют пространственно-частотные и ориентационные характеристики этих фрагментов. Здесь формируются группы нейронов, настроенные на разные ориентации и частоты, но направленные на один фрагмент. Они определяют его спектральный состав и связь с фрагментами иного спектрального состава.
На третьем уровне нейроны 19-го поля зрительной коры оценивают пространственное распределение возбуждения в нейронах предыдущего уровня и сопоставляют фрагменты изображения на основе их спектрального состава. Здесь также работают модули из нейронов разных свойств, обрабатывающих один участок зрительного поля, в результате чего выделяются отдельные компоненты фигур.
Наконец, четвертый уровень включает в себя обучающиеся нейроны, расположенные в нижневисочной и заднетеменной коре (НВК и ЗТК). Путем синтеза информации, поступающей от нервных механизмов предыдущего уровня, НВК обеспечивает опознание зрительного образа, ЗТК — его конкретизацию в пространстве.